| Министерство и образование науки Российской Федерации
Курсовая работа
По экологии
На тему: «Мутуализм как форма межвидовых контактов»
.
Санкт-Петербург
2011г.
Содержание. 2
Введение. 3
1.Общая характеристика взаимоотношений между организмами в биоценозе. 3
2.Мутуализм.. 3
3.Примеры мутуалистических взаимоотношений. 3
3.1.Примеры мутуализма, включающего поведенческие взаимосвязи. 3
3.2. Мутуализм, включающий разведение растений или животных. 3
3.3. Мутуализм при опылении. 3
3.4. Мутуализм с участием организмов, населяющих пищеварительный тракт. 3
3.5. Симбионты, живущие в тканях или клетках животных. 3
3.6. Мутуализм высших растении и грибов — микориза. 3
4. Эволюция субклеточных структур при симбиозе Мутуализм — возможный источник эволюции эукариот. 3
5. Модели мутуализма. 3
6. Некоторые общие черты жизнедеятельности мутуалистов. 3
Заключение. 3
Список литературы.. 3
Данная работа посвящена рассмотрению в природе такого типа взаимоотношения, как мутуализм.
Для меня эта тема в первую очередь интересна тем, что почти все живые существа в природе, так или иначе, вовлечены в этот тип взаимодействия, следовательно, мутуализм наиболее распространенный тип отношений между организмами. Некоторые типы взаимоотношений (рассмотренные ниже) могут со временем стать мутуалистическими отношениями, в то время как редко мутуализм может перейти в паразитизм, например.
В своей работе я рассмотрела мутуализм и его типы, примеры перехода от сотрудничества к симбиотическому мутуализму. Небольшую главу посвятила роли мутуализма в эволюции.
В конце представлены некоторые общие черты жизнедеятельности мутуалистов.
Начну свою работу с классификации и краткого описания этих взаимоотношении в биоценозе.
Каждый организм в биоценозе занимает определенное пространство, которое называют экологической нишей. Экологическая ниша
—
место вида в природе, преимущественно в биоценозе, включающее как положение его в пространстве, так и функциональную его роль в сообществе, отношение к абиотическим условиям существования (Хрусталев, Матишов, 1996). Важно подчеркнуть, что эта ниша не просто физическое пространство, занимаемое организмом, но и его место в сообществе, определяемое его экологическими функциями. Ю. Одум (1975) образно представил экологическую нишу как занятие, «профессию» организма в той системе видов, к которой он принадлежит, а его местообитание — это «адрес» вида.
Забиваем Сайты В ТОП КУВАЛДОЙ - Уникальные возможности от SeoHammer
Каждая ссылка анализируется по трем пакетам оценки: SEO, Трафик и SMM.
SeoHammer делает продвижение сайта прозрачным и простым занятием.
Ссылки, вечные ссылки, статьи, упоминания, пресс-релизы - используйте по максимуму потенциал SeoHammer для продвижения вашего сайта.
Что умеет делать SeoHammer
— Продвижение в один клик, интеллектуальный подбор запросов, покупка самых лучших ссылок с высокой степенью качества у лучших бирж ссылок.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз,
а первые результаты появляются уже в течение первых 7 дней.
Зарегистрироваться и Начать продвижение
Знание экологической ниши позволяет ответить на вопросы. как, где и чем питается вид, чьей добычей он является, каким образом и где он отдыхает и размножается (Дажо, 1975).Модель экологической ниши, предложенная Г. Е. Хатчинсоном, довольно проста: достаточно на ортогональных проекциях отложить значения интенсивности различных факторов, а из точек пределов толерантности восстановить перпендикуляры, то ограниченное ими пространство и будет соответствовать экологической нише данного вида (рис. 1). Экологическая ниша — это область комбинаций таких значений факторов среды, в пределах которой данный вид может существовать неограниченно долго.

Рисунок
1
- Модель экологической ниши (по Г.Е. Хатчинсону)
Например, для существования наземного растения достаточно определенного сочетания температуры и влажности, и в этом случае можно говорить о двумерной нише. Для морского животного уже необходимо кроме температуры еще как минимум два фактора — соленость и концентрация кислорода — тогда уже следует говорить о трехмерной нише (рис. 1),
и т. д. На самом деле этих факторов множество и ниша многомерна.
Экологическую нишу, определяемую только физиологическими особенностями организмов, называют фундаментальной, а ту, в пределах которой вид реально встречается в природе, — реализованной.
Реализованная ниша
— это та часть фундаментальной ниши, которую данный вид. популяция в состоянии «отстоять» в конкурентной борьбе. Конкуренция, по Ю. Одуму (1975,1986), — отрицательные взаимодействия двух организмов, стремящихся к одному и тому же (табл.1). Межвидовая конкуренция — это любое взаимодействие между популяциями, которое вредно сказывается на их росте и выживании. Конкуренция проявляется в виде борьбы видов за экологические ниши.
Классификация биотических взаимодействий популяций двух видов приведена в табл.1.
В таблице 1 «О» означает, что популяция не испытывает никакого влияния при взаимодействии видов; « + » — что она получает пользу от взаимодействия видов; «-» — что она испытывает отрицательное влияние такого взаимодействия.
| Классификация биотических взаимодействий популяций двух
видов (по Ю.Одуму,1986)
Таблица 1
| Тип
взаимодействия
|
Виды
|
Общий характер взаимодействия
|
| 1
|
2
|
| 1.нейтрализм
Сервис онлайн-записи на собственном Telegram-боте
Попробуйте сервис онлайн-записи VisitTime на основе вашего собственного Telegram-бота:
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно.
Зарегистрироваться в сервисе
|
0
|
0
|
На одна популяция не влияет на другую
|
| 2.конкуренция непосредственное взаимодействие
|
-
|
-
|
Прямое взаимное подавление обоих видов
|
| 3.конкуренция,взаимодействие из-за ресурсов
|
-
|
-
|
Непрямое подавление при дефиците внешнего ресурса
|
| 4.аменсализм
|
-
|
0
|
Популяция 2 подавляет популяцию 1, но сама не испытывает отрицательного воздействия
|
| 5.паразитизм
|
+
|
-
|
Популяция-паразит 1 состоит из меньших по величине особей, чем популяция 2
|
| 6.хищничество
|
+
|
-
|
Особи хищника 1 обычно крупнее, чем особи жертвы 2
|
| 7.комменсализм
|
+
|
0
|
Популяция1 , комменсал, получает пользу от объединения; популяции 2 это объединение безразлично
|
| 8.протокооперация
|
+
|
+
|
Взаимодействие благоприятно для обоих видов, но не обязательно
|
| 9.мутуализм
|
+
|
+
|
Взаимодействие благоприятно для обоих видов и обязательно
|
|
Не существует двух различных видов, занимающих одинаковые экологические ниши, но есть близкородственные виды, часто настолько сходные, что им требуется, по существу, одна и та же ниша. В этом случае, когда ниши частично перекрываются, возникает особо жесткая конкуренция, но в конечном итоге нишу занимает один вид. Явление экологического разобщения близкородственных (или сходных по иным признакам) видов получило название принципа конкурентного исключения, или принципа Гаузе, в честь ученого, доказавшего его существование экспериментально в 1934 г. (рис. 2).
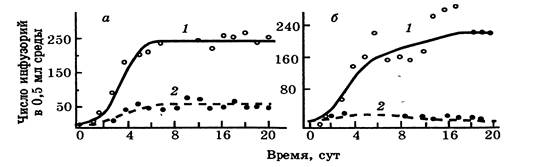
Рисунок 2 - Динамика популяций инфузорий Paramecium aurelia(1) и Paramecium caudatum (2),культивируемых при регулярном добавлении в среду одного и того же количества пищи: а-изолированные популяции каждого вида; б- совместно культивируемые популяции (по Гаузе,19)
Г.Ф. Гаузе экспериментально исследовал конкуренцию двух видов инфузорий: Paramecium candatum и Paramecium aurelia. Их культивировали раздельно и вместе, используя строго дозированную бактериальную пищу. При раздельном культивировании их численность росла по обычной S-образной кривой, при совместном — побеждали в конкурентной борьбе P. Aurelia (рис. 2). Поражение P. candatum объясняется тем, что она плохо переносила накопление в среде продуктов метаболизма бактерий и размножалась медленнее. Но при смене пищи, например при замене ее на дрожжи, побеждала уже P. candatum, так как в благоприятных для обоих видов условиях она имела преимущество за счет способности к более быстрому размножению и увеличению своей численности.
Межвидовая конкуренция за ресурсы может касаться пространства, пищи, биогенных веществ и т. п. Именно уменьшение ресурсов приводит к ситуациям, когда мы имеем дело лишь с отрицательными взаимодействиями. Результатом межвидовой конкуренции может быть либо взаимное приспособление двух видов, либо популяция одного вида замещается популяцией другого вида, а первый вынужден переселиться на другое место или перейти на другую пищу. Если виды живут в разных местах, то говорят, что они занимают разные экологические ниши, если же они живут в одном месте, но потребляют разную пищу, то говорят об их несколько различающихся экологических нишах. Процесс разделения популяциями видов пространства и ресурсов называется дифференциацией экологических ниши (рис. 3).

Рисунок 3 - Распределение копытных зверей по ярусам питания в африканской саванне: 1-жираф;2-антилопа геренук;3-антилопа дик-дик; 4-носорог;5-слон;6-зебра;7-гну;8-газель Гранта; 9-антилопа бубал(по де ла Фуэнте,1972)
Главный результат дифференциации ниш — снижение конкуренции. Например, тенелюбивые растения не конкурируют со светолюбивыми, менее остра конкуренция за ресурсы, численность доминирующего вида, например, регулируется хищником, и т. п. Иными словами, есть множество обстоятельств, при которых разные виды-антагонисты могут сосуществовать. И тем не менее это отрицательные взаимодействия, поскольку взаимовлияние видов остается и не позволяет полностью раскрыть свои возможности каждому из них.
Нейтрализм
— это такая форма биотических взаимоотношений. когда сожительство двух видов на одной территории не влечет за собой ни положительных, ни отрицательных последствий для них. В этом случае вилы не связаны непосредственно друге другом и даже не контактируют между собой. Например, белки и лоси, обезьяны и слоны и т. п. Отношения нейтрализма характерны для богатых видами сообществ.
Аменсализм
— это биотические отношения, при которых происходит торможение роста одного вида (аменсала) продуктами выделения другого. Такие отношения обычно относят к прямой конкуренции и называют антибиозом. Наиболее хорошо они изучены у растений, которые применяют различные ядовитые вещества в борьбе с конкурентами за ресурсы, и данное явление называют аллелопатия.
Аменсализм весьма распространен в водной среде. Например, сине-зеленые водоросли, вызывая цветение воды, тем самым отравляют водную фауну, а иногда даже скот, который приходит на водопой. Аналогичные «способности» проявляют и другие водоросли. Они выделяют пептиды, хинон, антибиотики и другие вещества, которые ядовиты даже в малых дозах. Называют эти яды эктокринными веществами.
Хищничество и паразитизм
: отношения хищник — жертва и паразит — хозяин являются результатом прямых пищевых связей, которые для одного из партнеров имеют отрицательные последствия, а для другого — положительные. Все варианты пищевых экологических связей можно отнести к этих типам взаимодействия (в том числе и корова, поедающая траву). Любой гетеротрофный организм в сообществе существует за счет поедания другого гетеро- или автотрофа.
Хищниками называют животных, питающихся другими животными, которых они ловят и умервщляют. Для хищников характерно охотничье поведение. Изобилие насекомых, их малые размеры и легкодоступность превращают деятельность плотоядных хищников, обычно птиц, в простое «собирательство» добычи, подобно тому как собирают семена, зерна птицы, питающиеся ими. Насекомоядные хищники по способу овладения пищей приближаются к пастьбе травоядных животных. Некоторые птицы могут питаться и насекомыми и семенами.
Паразитизм — это такая форма пищевой связи между видами, когда организм-потребитель (консумент) использует тело живого хозяина не только как источник пищи, но и как место своего обитания (постоянного или временного). Паразиты намного мельче своего хозяина. Паразитические отношения имеют насекомые-вредители и растения, кровососущие насекомые и животные, и т. п. Насекомые-паразиты часто бывают разносчиками эпидемий: вши — тифа, клещи — энцефалита, и др.
В природе существуют системы, состоящие из одного вида и нескольких других видов, являющихся по отношению к нему паразитами. Это так называемые паразитарные комплексы.
Например, чтобы успешно бороться с вредителями культурных растений, необходимо изучать состав и плотность комплекса, закономерности его роста, и т. п.
Хищничество и паразитизм — это пример взаимодействия двух популяций, отрицательно сказывающееся на росте и выживании одной из них (см. табл. 1, п. 5, 6). Подобные популяции развиваются, т. с. эволюционируют, синхронно, и по мере длительности их взаимодействия коэволюция может привести к снижению степени отрицательного взаимодействия или устранить его вообще, поскольку сильное подавление популяции жертвы или хозяина популяцией хищника или паразита может привести к уничтожению одной из них или обеих.
На рис. 4 приводится пример эволюции гомеостаза двух насекомых в системе «хозяин-паразит», которые помещались в клетку, состоящую из 30 пластиковых камер, соединенных друг с другом трубочками, замедлявшими расселение паразита. На Рис. 4 а
видны резкие подъемы и спады плотности популяций, так как в этом случае дикие особи недавно посажены вместе. На рис. 4 б популяции взяты из колоний, в которых они просуществовали совместно в течение двух лет и здесь уже отмечается более стабильное равновесие, резкие спады отсутствуют, так как у хозяина появляется адаптивная устойчивость, о чем свидетельствует сильное снижение рождаемости у паразита.
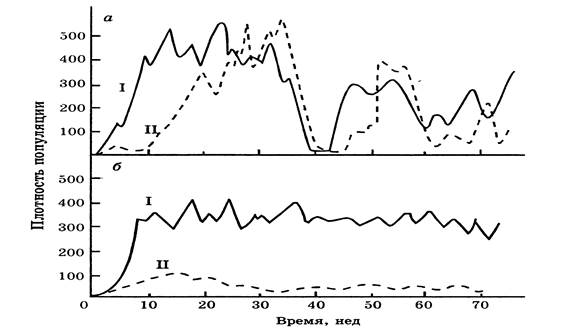
Рисунок 4 - Эволюция гомеостаза в системе "хозяин-паразит"(хозяин комнатная муха Musca domestica (I),паразит - оса Nasonia vitropennis(II)(по Ю.Одуму):
а
- недавно объединенные популяции, б
- популяции взятые из колоний, в которых оба вида существовали два года
Итак, наиболее жесткая конкуренция проявляется тогда, когда контакт между популяциями установлен недавно, например, вследствие изменений, произошедших в экосистеме под влиянием деятельности человека. Именно поэтому непродуманное вмешательство человека в структуру биоценоза нередко приводит к эпидемическим вспышкам.
Таким образом, при длительном контакте паразитов и хищников с их жертвами, влияние на них весьма умеренно, нейтрально или даже благоприятно, а наибольшее повреждающее действие оказывают новые паразиты и хищники. Отсюда вывод: «...необходимо избегать создания новых отрицательных взаимодействий, а если они возникли, стараться по возможности сдерживать их» (Ю. Одум, 1975).
К положительным видам взаимодействия Ю. Одум относит комменсализм, кооперацию и мутуализм (см. табл. 1). Многие экологи считают, что в стабильных экосистемах отрицательные и положительные взаимодействия должны находиться в равновесии.
Комменсализм, кооперацию и мутуализм можно рассматривать как стадии последовательного совершенствования положительных взаимодействий в ходе эволюции.
Комменсализм
— это наиболее простой тип положительных взаимодействий (см. табл. 4.). Комменсалы — организмы. которые поселяются в жилищах других организмов, не причиняя им зла и не принося добра. Для тех животных, у которых они «квартируют», комменсалы безразличны. В океанах и морях в каждой раковине есть организмы, которые получают там укрытие, но они абсолютно безобидны для «владельца» этой раковины.
Ниже, более подробно будет рассмотрен непосредственно такой вид взаимодействия как мутуализм.
Мутуализмом называют взаимодействия между парами видов, приносящие обоюдную пользу, т. е. в популяции каждого из этих видов (мутуалистов) особи растут и (или) выживают и (или) размножаются с большим успехом в присутствии особей другого вида. Преимущества могут быть разные. Чаще всего они заключаются в том, что по крайней мере один из партнеров использует другого в качестве пищевого ресурса, тогда как другой получает защиту от врагов или благоприятные для роста и размножения условия. В других случаях вид, выигрывающий в пище, освобождает партнера от паразитов (например, рыбы-чистильщики). Несмотря на преимущества, получаемые каждым из партнеров, при рассмотрении мутуализма важно помнить, что речь отнюдь не идет о дружеской взаимопомощи. Каждый партнер, по существу, действует «эгоистично» и выгодные отношения возникают просто потому, что получаемая им польза перевешивает требуемые затраты.
В ранних работах по экологии мутуализм обычно недооценивали или вообще упускали из виду. Тем не менее, это явление распространено очень широко. От него зависит образование значительной доли биомассы планеты — например, доминантами лугов и лесов (корни растений тесно связаны с грибами —микориза), а также корралами (в полипах находятся одноклеточные водоросли).
Чрезвычайно распространен в природе и, вероятно, имеет не менее важное значение для определения природы популяций и сообществ, чем конкуренция, паразитизм и т. д.. Положительные взаимодействия формировались в данной последовательности в ходе эволюции: комменсализм (преимущества имеет одна популяция), протокооперация (пользу получают обе популяции) и мутуализм (пользу получают обе популяции, причем они полностью зависят друг от друга).
Широкое распространение выдвинутое Дарвином представления о «выживании наиболее приспособленных» как важном механизме естественного отбора послужило причиной того, что внимание исследователей преимущественно сосредоточивалось на различных проявлениях конкуренции в природе. В результате значение кооперации между видами оказалось недооцененным. Во всяком случае, положительные взаимодействия изучены количественно далеко не столь подробно, как отрицательные взаимодействия. По-видимому, есть все основания допустить, что отрицательные и положительные отношения между популяциями в экосистемах, которые достигают некоторого стабильного состояния, в конце концов уравновешивают друг друга. Наиболее простой тип положительных взаимодействий, который, видимо, представляет собой и первый шаг на пути их развития, — комменсализм. Он характерен, в частности, для взаимоотношений между прикрепленными растениями и животными, с одной стороны, и подвижными организмами — с другой. Многие комменсалы используют в качестве хозяина один определенный вид, другие же — разные виды.
Лишь один шаг отделяет комменсализм от такой ситуации, при которой оба организма получают преимущества от объединения или какого-либо иного взаимодействия друг с другом; в этом случае мы имеем дело с протокооперацией. Много работал в этой области и писал по этому вопросу покойный У. Олли (1938, 1951). Он полагал, что зачатки кооперации между видами встречаются в природе повсюду. Много случаев кооперации он установил сам и экспериментально доказал ее пользу для обоих видов.
Следующий этап—развитие зависимости обеих популяций друг от друга. Такие случаи были названы мутуализмом.
Мутуализм включает разнообразные формы сотрудничества – от облигатного (симметричного или асимметричного), при нарушении которого гибнут оба или один сотрудничающий партнер, до факультативного, которое помогает выживать партнерам, но не является для них обязательным (так называемая протокооперация).
Рассмотрим основные варианты мутуализма. Объединение часто происходит между весьма разными организмами с сильно различающимися потребностями, и это, по-видимому, совершенно естественно. (Между организмами со сходными потребностями чаще всего возникают отрицательные взаимодействия.) Наиболее важные мутуалистические системы возникают между автотрофами и гетеротрофами. Это и не удивительно, поскольку эти два компонента экосистемы должны в конце концов достигнуть некоторого равновесного симбиоза. Однако общая взаимозависимость, часто наблюдаемая в сообществах, не относится к категории явлений, которые мы обозначили как мутуализм; последний имеет место лишь тогда, когда какой-либо определенный гетеротроф становится полностью зависимым от некоторого определенного автотрофа в отношении пищи, а существование последнего зависит от защиты, обмена веществ и других жизненно важных функций гетеротрофа. Как отмечалось ранее, мутуалистические отношения, по-видимому, замещают паразитизм в ходе созревания экосистемы; они, вероятно, особенно важны в тех случаях, когда лимитированы некоторые ресурсы среды (например, на почве с низким плодородием); в такой ситуации взаимная кооперация может обеспечить весьма существенное селективное преимущество.
Сначала рассматривается ряд примеров мутуалистических связей между особями разных видов, начинающийся факультативным мутуализмом (каждый партнер или симбионт получает пользу от другого, но не зависит от него), продолжающийся взаимодействиями, облигатными для одного, но факультативными для другого партнера, и наконец, завершающийся отношениями, облигатными для обоих. Затем обсуждаются примеры, в которых один вид населяет поверхность или полость тела другого, а также случаи, когда один партнер (эндобионт) обитает внутри клеток другого, причем оба организма тесно связаны физиологически (рис. 5). организма тесно связаны физиологически (рис. 5).
Классификация преимуществ, получаемых партнерами при различных видах мутуализма, требующих сложных поведенческих связей, — непростая задача. Их разнообразие иллюстрируют следующие примеры.
3.1.1. Медоуказчик и медоед
Африканская птица -медоуказчик замечательным образом связана с млекопитающим, капским медоедом. Она разыскивает пчелиные гнезда и приводит -к ним партнера. Млекопитающее вскрывает гнездо и поедает мед и личинок пчел, а медоуказчик питается остатками его трапезы. Птица может разыскивать пчелиные гнезда, но не способна их вскрыть; медоед, наоборот,
легко их вскрывает, «о находит с трудом. Взаимосвязанное поведение приносит животным обоюдную -пользу.
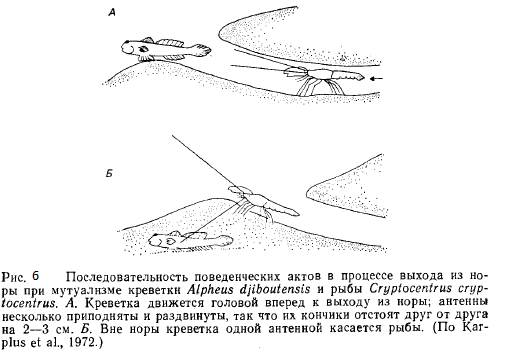
3.1.2. Креветки и бычки
Креветки рода Alphaeus выкапывают норы, которые бычки Cryptocentrus используют как убежища в среде, где иначе трудно укрыться от врагов. Креветка почти ничего -не видит и, когда покидает -нору, с помощью своих антенн постоянно поддерживает контакт с рыбой, получая таким образом предупреждения о любых изменениях обстановки (рис. 6). Бычок пользуется местообитанием с донными осадками, содержащими обильный корм, а креветка — зрительной системой партнера, позволяющей ей, не подвергаясь опасности, на короткое время покидать нору и питаться снаружи от нее (Frickc, 1975).
3.1.3. Рыба-клоун и актиния
Разнообразные типы мутуалистических поведенческих отношений встречаются у обитателей тропических коралловых рифов (мутуалистами являются сами кораллы — см. ниже). Рыба-клоун Atnphiprion живет рядом с актиниями (например, Physobrachia, Radianthus) и прячется среди их щупалец от врагов. Внутри актинии, рыба покрывается защитной слизью, предохраняющей от действия стрекающих нематоцист (эта слизь нужна самой актинии, чтобы предотвратить выстреливания нематоцист при соприкосновении соседних щупалец). Рыба, безусловно, получает защиту, но актинии это сотрудничество также полезно, поскольку рыба-клоун нападает на приближающихся рыб, включая и тех, которые обычно питаются актиниями (Fricke, 1975).
3.1.4. Рыбы-чистильщики и их клиенты
Еще одна явно мутуалистическая связь существует между рыбами, называемыми «чистильщиками», и их «клиентами». Чистильщики поедают эктопаразитов, бактерий и отмершие ткани с поверхности тела клиента. Различают сорок пять видов таких рыб. Они часто держатся на постоянном участке, тогда как их клиенты периодически посещают эти «пункты чистки»; причем тем чаше, чем больше на них паразитов. У обоих партнеров характерное поведение, позволяющее им узнавать друг друга и возникшее, по-видимому, в ходе тесной коэволюции. Чистильщики получают источник корма, а клиенты освобождаются от вредных объектов. В эксперименте на Багамских островах (Limbaugh, 1961) с отдельных участков рифа удалили всех рыб-чистильщиков. У рыб- клиентов развивались кожные заболевания и за две недели численность их популяций снижалась. Сходный эксперимент на Гавайских островах дал отрицательный результат, тем не менее вполне вероятно, что отношения чистильщик—клиент часто действительно мутуалистические. Обнаружено также, что чистильщиками рыб являются б видов креветок.
3.1.5. Муравьи и акация
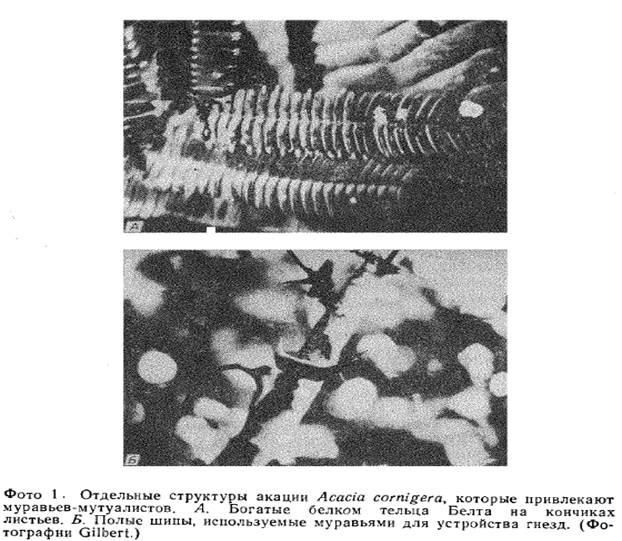
Еще одно такое мутуалистическое взаимодействие типа чистильщик—клиент описано (Janzen, 1967) в Центральной Америке у одной из акаций (Acacia cornigera) с муравьями Pseudomyrrnex ferruginea. В полых шипах этого растения муравьи строят гнезда, а листочки перистых листьев дерева несут на концах богатые белком тельца Белта (фото 1), которые муравьи собирают и используют в пищу; кроме того, на листьях муравьев привлекают выделяющие сахар нектарники. Со своей стороны, насекомые защищают эти небольшие светолюбивые деревца от конкурентов, активно «обстригая» чужие побеги, проникающие в их крону, а также защищают растение от фитофагов.
Этот пример мутуализма позволяет по-новому взглянуть и на возможную роль других растительных структур. Нектарники на вегетативных частях встречаются по меньшей мере у видов 39 семейств, населяющих многие экосистемы по всему свету. Если их назначение на цветках легко объяснимо необходимостью привлечения опылителей, то в данном случае оно менее понятно. Они, безусловно, привлекают муравьев, иногда в огромном количестве, но интерпретируя это явление, «протекционисты» (считающие, что муравьи защищают растения от фитофагов) расходятся с «эксплуатационистами», видящими в муравьях не больше пользы для растений, чем в блохах для собак (скептическая точка зрения, вполне допустимая в научных исследованиях). Обзор этой проблемы сделан Бентли (Bentley, 1977). Такого рода расхождения в толковании могут быть устранены в ходе тщательно разработанных и контролируемых экспериментах типа проведенного с растением Helianthella quinqucnervis (Inouye, Taylor, 1979), имеющего расположенные вне цветков нектарники. Этот вид может сильно страдать от мух-терфритид, личинки которых, если с растения удалить всех муравьев, повреждают иногда более 85% развивающихся семян. Контрольные экземпляры с муравьями от такого ущерба почти полностью избавлены.
3.2.1. Человек как мутуалист культурных растений
и домашних животных
Наиболее впечатляющие примеры мутуализма мы находим в сельском хозяйстве. Численность растений пшеницы, ячменя, овса, кукурузы и риса, а также площади, занятые под них, во много раз больше, чем были бы без специального культивирования. Рост народонаселения со времен охотников и собирателей может служить мерой пользы, которую человек извлекает из этого взаимодействия. Нельзя проверить этого экспериментально, но легко представить себе, к каким последствиям для популяции, например, риса во всем мире привело бы исчезновение человека или как отразилось бы на человечестве уничтожение всех посевов риса. Стоит отметить, что человек разводит и такие растения, как кофе, табак и опиумный мак, содержащие сильнодействующие защитные вещества (кофеин, никотин и т.д.). Очевидно, что растения получают при этом выгоду, но получат ли от них пользу люди, зависит от способа применения этих алкалоидов. Одомашнивание крупного рогатого скота, овец и других млекопитающих также связано с мутуализмом — без человека множество таких животных быстро погибнет, а у людей без домашних животных сильно изменится рацион питания.
Сходные мутуалистические отношения развились в сообществах термитов и муравьев.
3.2.2. «Разведение» гусениц муравьями
Классический пример мутуализма, включающего разведение одним животным другого, — взаимодействие между бабочкой голубянкой Lycacna avion и муравьями. Бабочка откладывает яйца на бутоны тимьяна, и ее личинки очень похожи на цветки, которыми они питаются. Спустя приблизительно 20 сут (после третьей линьки) гусеница покидает растение и начинает целенаправленно перемещаться— растениями она больше никогда не питается. Когда личинку находит муравей, в ответ на его поглаживание она выделяет из медоносной железы капельки жидкости, которые муравей выпивает. После сложной последовательности взаимных сигнальных действий он переносит гусеницу в свою подземную колонию, где она находится около 11 месяцев. Часть этого времени она пребывает в зимней спячке или в состоянии куколки, но когда бывает активна, то питается молодыми личинками муравьев. В июне, завершив личиночное
развитие, бабочка покидает муравейник.
Жизненный цикл этой голубянки столь причудлив, что может показаться невероятным, если не знать, что он является лишь частью сложного эволюционного ряда взаимодействий между муравьями и многими представителями семейства Lycaenidae, к которому она принадлежит ( Ford, 1945). Другие бабочки из этого семейства также «пасутся» муравьями (хотя не всегда внутри их колоний), и гусеницы у большинства его видов имеют медоносные железы, которые «выдаивают» муравьи. Практически так же они «пасут» и «доят» многие виды равнокрылых, получая от них сахаристые «выделения. Муравьи защищают этих насекомых от хищников и паразитоидов, аналогично тому как в обмен на секрет внецветковых нектарников, они по-видимому, охраняют соответствующие растения от фитофагов (Strong et al., 1984).
В Европе бабочка L. avion — редкий, находящийся под угрозой исчезновения вид. Недавно она исчезла в Великобритании. Сложный цикл развития делает ее полностью зависимой от присутствия цветущего тимьяна, а затем — личинок муравьев. Партнерами, «по-видимому, могут быть только муравьи двух видов (Myrmica scabrinoides и М. laevonoides), а луга на известняках, служащие местообитанием как для тимьяна, так и для этих муравьев, быстро исчезают. Жизненный цикл этого вида является, таким образом, классическим примером «слишком узкой специализации», при которой особенно велика опасность вымирания.
3.2.3. Разведение грибов жуками
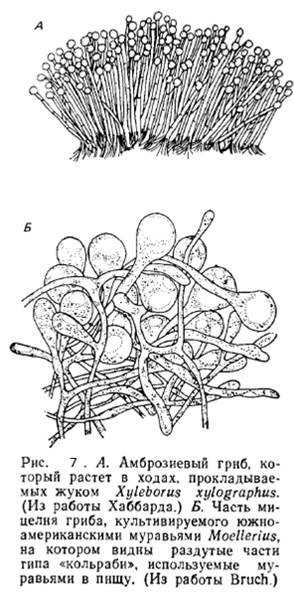 Большая часть тканей растений, включая древесину, недоступна животным в качестве непосредственного источника пищи, потому что у них нет ферментов для переваривания целлюлозы и лигнина . Однако такие ферменты есть у многих грибов, и питающиеся ими животные, косвенно получают доступ к богатому энергией корму. В ряде случаев между ними Большая часть тканей растений, включая древесину, недоступна животным в качестве непосредственного источника пищи, потому что у них нет ферментов для переваривания целлюлозы и лигнина . Однако такие ферменты есть у многих грибов, и питающиеся ими животные, косвенно получают доступ к богатому энергией корму. В ряде случаев между ними
и грибами - редуцентами возникают весьма специализированные мутуалистические отношения. Жуки-короеды (семейство Scolytidae) прокладывают в мертвой или отмирающей древесине глубокие ходы, в которых поселяются специфичные для данного вида жуков грибы, постоянно выедаемые личинками. Эти насекомые, вероятно, переносят «посевной материал» грибов в своем пищеварительном тракте, хотя у некоторых видов на голове обнаружены специальные щетки из волосков, собирающие споры. Грибы служат жукам пищей и используют их при заселении новых ходов в древесине—отношения вполне мутуалистические (рис. 7).
3.2.4. Разведение грибов муравьями
У некоторых видов муравьев (и термитов) наблюдается еще более тонкая система разведения грибов. Самым замечательным примером можно считать здесь муравьев рода Atta. Они выкапывают в почве полости объемом два — три литра. Грибы культивируются в них на листьях, срезаемых с ближайших растений, причем вся колония муравьев иногда полностью зависит от этого источника пищи. И в данном случае грибы выигрывают, так как не только поедаются, но и расселяются. Готовая к размножению самка, покидая старую «колонию, чтобы основать новую, заполняет «посевным материалом» гриба особый карман в своей глотке.
Цветки, опыляемые насекомыми, бывают «универсалами» и специалистами». — Выгоды специализации. — Мутуализм при опылении инжира бластофагами.
Большинство животных, участвующих в опылении цветков, в качестве «вознаграждения» получают нектар или пыльцу. Нектар, по-видимому, служит растению только для привлечения опылителей и образование его представляет собой в сущности расход веществ, в основном углеводов, которые оно могло бы использовать само для роста или других целен. Однако эти траты компенсируются пользой от опыления. (Столь же «расточительны» многие виды растений, образуя плоды, которые привлекают животных и служат им кормом; выгода заключается в расселении содержащихся в плодах семян).
Опыление — это процесс переноса пыльцы с пыльника одного цветка на рыльце другого, иногда на том же растении. Множество цветковых видов, особенно однолетников, в норме являются самоопыляемыми, т. е. способны к самооплодотворению и не нуждаются в переносчике пыльцы. Прочие тесно связаны с опылителями, обеспечивающими перекрестное опыление и перекрестное оплодотворение. По-видимому, эволюция специализированных цветков и способов привлечения к ним животных вызвана тем, что внутривидовой аутбридинг и(или) избегание межвидового аутбридинга дают существенные адаптивные преимущества. Мы не будем обсуждать возможные выгоды, даваемые некоторым растениям за счет развития систем аутбридинга (по-видимому, приводящего как к генетическим, так и к энергетическим затратам). Как и в отношении самого полового размножения, причины эволюции аутбридинга еще не понятны ( Williams, 1975; Maynard, Smith, 1978). Однако они были столь серьезными, что обусловили значительную долю разнообразия в царстве растений и возникновение некоторых из наиболее тонких мутуалистнческих связей.
Опылителями цветковых растений являются и теплокровные, в том числе колибри, летучие мыши и даже мелкие грызуны и сумчатые (фото 13), однако, без сомнения, основные опылители— это насекомые (Grant, 1963).
В простейших случаях опыления насекомыми нектар или пыльца имеются в изобилии и легко доступны для самых разнообразных посетителей. Примером служит ежевика (Rubus fruticosus), которая выделяет нектар в таком количестве, что местами его даже слишком много для многочисленных питающихся им видов. У других растении различным образом специализированные структуры цветка защищают нектар от всех посетителей, кроме нескольких видов насекомых, имеющих соответственно устроенные ротовые органы. В семействе лютиковых прослеживается последовательная эволюция формы нектарников — от открытых у Ranunculus ficaria через простые крышечки, защищающие нектароносную зону лепестков, к сложным заполненным нектаром нектарникам-шпорцам у Aquilegia. Специализация цветков ограничивает разнообразие насекомых,
способных собирать нектар и, следовательно, играть роль опылителей. Это приводит к трем основным последствиям: 1) опылители, как правило, посещают экземпляры только одного вида (возможно, это связано с формированием у насекомого «образа искомого»), что повышает вероятность внутривидового аутбридинга; 2) на рыльцах других видов теряется меньшее количество пыльцы данного растения; 3) в результате научения или эво-
люции у насекомых могут развиваться специальные навыки, делающие их более эффективными сборщиками пищи по сравнению с опылителями-универсалами.
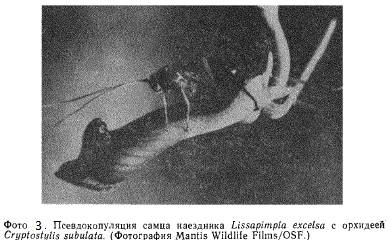
Процесс опыления у орхидных дает примеры наиболее тесных, по -видимому, возникших в результате коэволюции, мутуалистических взаимоотношений (фото 3), которые по крайней мере для одного из партнеров облигатны. Другие тонко организованные их варианты могли и не требовать коэволюции (взаимного отбора каждого из партнеров другим).
 Например, разные Например, разные
представители рода ваточник (Asclepias) опыляются шмелями
одного и того же вида, но их цветки устроены так, что пыльцевая масса пристает к различным частям тела насекомого и попадает затем только на рыльце растения соответствующего вида. Это препятствует межвидовому перекрестному опылению, однако и здесь можно видеть, что, хотя шмель остается неспециализированным опылителем, виды ваточника вынуждены специализироваться.
У большинства растений цветение происходит в определенный сезон, и это строго ограничивает степень специализации опылителя. Его жизненный цикл может стать полностью зависимым от определенных цветков как источника пищи только в том случае, если совпадает со временем их распускания. Это возможно для насекомых, использующих цветки в течение короткого периода (например, только для питания, как у чешуекрылых). Долгоживущие опылители, например летучие мыши и грызуны или пчелы с их долговременными колониями, вероятно все же до определенной степени являются универсалами, поочередно использующими в течение сезона цветы разных видов или, если нектар недоступен, переходящими на другие виды корма.
Замечательный и совершенно особый тип облигатного для обоих партнеров мутуализма известен между инжиром (Ficus, семейство Могасеае) и его опылителем, бластофагой из перепончатокрылых (Wlebes, 1979; Janzen, 1979).
Цветки в этом случае находятся в полости, образуемой разросшимся ложем соцветия (мясистыми тканями стебля), причем мужские и женские—на разных частях растения. Женские цветки бывают двух типов — длинно- и короткостолбиковые. Самки насекомых проникают в соцветия через их апикальное отверстие и откладывают яйца в завязи с коротким столбиком (для длинного столбика их яйцеклад слишком короток), становящиеся галлами, где личинки бластофаги завершают свое развитие. Для каждого вида инжира известен более или менее специфичный вид насекомых-партнеров. Самцы проделывают отверстие в соцветии, по которому за ними выходят наружу самки, но только после того, как посетят мужские цветки и соберут с них пыльцу. Пыльца переносится в специальных сумках: установлено, что когда самка проникает в новое соцветие, она вытряхивает из них пыльцу и помешает ее на рыльца длинностолбиковых завязей, в которых после опыления развиваются семена (фото 4).
 Такая строго определенная последовательность событий Такая строго определенная последовательность событий
должна была выработаться в условиях обоюдного давления
естественного отбора, когда каждый этап эволюции соцветия инжира вызывал ответное изменение в поведении бластофаг (Wiebes, 1982). Насекомое стало не только опылителем, но и паразитом. Эволюция этой сложной экосистемы пошла еще дальше. С большинством видов инжира связано два разных вида перепончатокрылых, каждый из которых специализирован на определенном типе цветков внутри соцветия (кроме того, обычно существует два вида паразитических перепончатокрылых, специализирующихся на этих двух видах-опылителях). Часто наблюдаются представители этого же семейства насекомых, которые откладывают яйца в женские цветки инжира, но не играют никакой роли в процессе опыления. Их нельзя считать мутуалистами, скорее они напоминают кукушку, подкидывающую яйца в гнезда, «свитые» инжиром, но никак не расплачивающуюся за услуги.
Этот тип взаимодействий с участием галлообразователей, обычных или «гнездовых» паразитов неоднократно встречается у различных диких видов инжира. Такая ситуация является одним из классических примеров коэволюции и параллельной эволюции мутуализма и паразитизма. (Отметим, что культурный инжир размножается партеногенетически и для образования плодов не требует опыления. Таким образом, он не зависит от бластофаг).
Пищеварительный тракт как аналог ферментера.
В большинстве обсуждавшихся до сих пор случаев мутуализм зависел от поведения участвующих во взаимодействии животных. Оно включало поиск партнера и обычно (но не всегда) пищевое вознаграждение. При таком мутуализме оба партнера являются эволюционно продвинутыми видами и каждый из них проводит значительную часть своей жизни самостоятельно.
Во многих других случаях один из партнеров — одноклеточный эукариот или прокариот, тесно связанный с жизнедеятельностью многоклеточного партнера и как бы являющийся его более или менее постоянной частью. Яркие примеры такого взаимодействия дает нам микробоценоз пищеварительного тракта.
У большинства животных этот тракт представляет собой микробный микрокосм. У многих растительноядных животных его микрофлора играет главную роль в переваривании целлюлозы и, возможно, также в синтезе витаминов. Здесь поддерживается более или менее постоянный приток субстратов в виде съеденной, пережеванной и частично гомогенизированной животными пищи.
В пищеварительном тракте регулируется рН субстрата, контролируется его аэрация и перемешивание (а у теплокровных —и температура). Отходы постоянно удаляются, благодаря чему предотвращается загрязнение. Такая система очень похожа на непрерывно действующий ферментер, используемый, например, в промышленном производстве пива, «когда пивовар регулирует тот же самый набор условий среды.
3.4.1. Экосистема рубца
Флора и фауна рубца, разлагающие целлюлозу. — В рубце присутствует полностью сформированное сообщество.
Желудок жвачных животных (к ним относятся например, олени, крупный рогатый скот и антилопы) состоит из четырех отделов, и проглоченная пища сначала попадает в тот из них, который называется сеткой. Первое пережевывание приводит к измельчению пищи до частиц объемом 1 — 1000 мкл, причем отдельные из них могут достигать в длину 10 см. Из сетки в следующий отдел желудка, книжку, могут проходить только
частицы объемом не более 5 мкл; более крупные животное отрыгивает и снова пережевывает (процесс непрерывного «жевания жвачки»). Рубец населен многочисленными бактериями (1010
—1011
в 1 мл) и простейшими (105
— 106
в 1 мл); рН среды в нем регулируется животным за счет выделения слюнными железами секрета, содержащего 100—140 мМ бикарбоната и
10—50 мМ фосфата. Таким образом, непрерывный приток субстратов и контроль условий его сбраживания микроорганизмами обеспечивается самим хозяином, а продукты микробной ферментации являются для него основным источником питания (рис. 9).
Бактериальное сообщество рубца почти полностью состоит
из облигатных анаэробов — многие эти виды под воздействием
кислорода мгновенно гибнут. Для них необходимы углеводы, а большинство нуждается также в уксусной, изомасляной, изовалериановой,
2-метилмасляной кислотах и аммиаке; многие из этих веществ поставляются другими населяющими рубец бактериями. Флора рубца отнюдь не является смесью микробов-универсалов. У многих из них специализированный обмен веществ, ограничивающий их распространение только этой уникальной средой.
Основными компонентами пищи жвачных являются целлюлоза и другие растительные волокна, но сами животные лишены ферментов, способных разлагать этот материал. Поэтому целлюлолитическая активность микрофлоры рубца имеет для них важнейшее значение. Bacteroides sucoinogenes плотно прилипает к растительным волокнам и расщепляет целлюлозу; Ruminococcus переваривает целлюлозу, целлобиозу и ксилозу; другие виды, например Clostridium locheadii, могут разрушать не только
целлюлозу, но и крахмал; некоторые узкие (пищевые специалисты, в частности Methanobacterium ruminantium, в качестве источника энергии способны использовать лишь водород или формиат: для Bacteroides amylophilus субстратом служит только крахмал и его производные.

В рубце эти виды занимают специализированные «метаболические ниши».
Фауна желудочных простейших также представляет собой сложное сочетание специализированных форм. Большинство из них — инфузории: равноресничные и энтодиниоморфы (последняя группа известна только из рубца; (фото 5). Свободноживущие простейшие рубца обитают в среде с постоянными условиями, где они, вероятно, интенсивно 'конкурируют с огромным числом сопутствующих микробов. По морфологическому
разнообразию и сложности эти сообщества сравнимы с тропическими экосистемами, в которых высокие продуктивность и видовое разнообразие поддерживаются благоприятными условиями среды (Hungate, 1975).

Только очень немногие простейшие способны переваривать целлюлозу (и даже они это делают с помощью своих собственных симбионтов-бактерий). Многие простейшие поедают бактерий, и если первых удалить, численность вторых увеличится. С другой стороны, многие энтодиниоморфы — хищники, питающиеся другими простейшими. Таким образом конкуренция, хищичество, мутуалиам и передача вещества по пищевым цепям, свойственные наземным и водным экосистемам, происходят и в микрокосме рубца. На самом деле, ситуация еще сложнее, поскольку его микрофлора у разных видов жвачных неодинакова. Кроме того, она может сильно измениться, если внезапно изменится состав пищи.
Микроорганизмы рубца постоянно размножаются и одновременно сокращают численность .по мере того как его содержимое переходит в кишечник. Дальнейшее переваривание пищи, в том числе и некоторых микробов происходит в кишечнике за счет собственных ферментов жвачного. Основные продукты пищеварения в рубце — летучие жирные кислоты (уксусная, пропионовая, масляная), аммиак, двуокись углерода и метан. Жирные кислоты всасываются и служат жвачному основным источником углеродного питания. Особенно важна пропионовая кислота,
единственная, которая может быть превращена этими животными в углеводы и незаменима для их обмена -веществ, особенно в период лактации.
Мутуалистический характер связи жвачных с микрофлорой рубца очевиден: микробы получают постоянный источник пищи и довольно стабильные условия, а животное — доступные для переваривания вещества из корма, который не может быть переработан с помощью его собственных ферментов.
3.4.2. Экосистема пищеварительного тракта термитов
Расщепление целлюлозы в кишечнике термитов ведут в основном простейшие. — В кишечнике термитов фиксируется молекулярный азот.
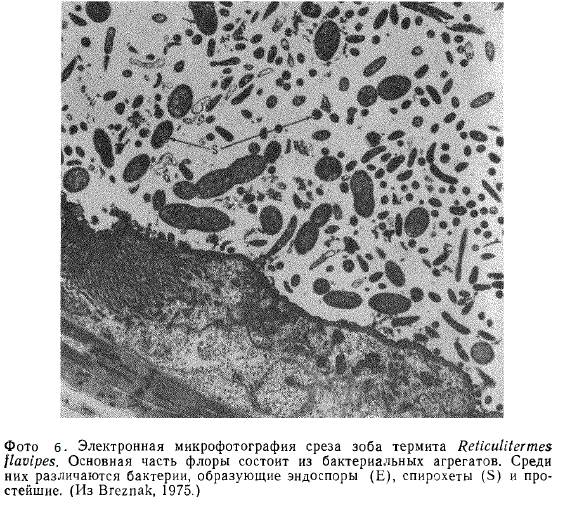
Термиты — это колониальные общественные насекомые (отряд Isoptera). Предположительно более продвинутые из них — представители группы Macrotermitineae разводят грибы. Термиты из «более отсталых» групп питаются непосредственно древесиной: целлюлоза, гемицеллюлоза и лигнин перевариваются в их пищеварительном тракте. Под воздействием
собственных ферментов насекомых предварительное расщепление происходит в их передней и средней кишке, но основная масса пищи поступает в расширенную часть сегментированной слепой кишки, представляющую собой место для микробной ферментации (фото 6).
Термиты поедают собственные экскременты и, таким образом, пищевой материал по меньшей мере дважды проходит через их кишечник, а микробы, размножившиеся при первом прохождении, во второй раз могут быть переварены сами. Основная группа организмов -в кишечнике термитов— это простейшие, а именно анаэробные жгутиконосцы,
например, Trichomonas termopsidis и представители уникальных
родов, обнаруженных только у термитов и у нескольких питающихся древесиной видов тараканов (Cryptocercus). Имеются и бактерии, но именно простейшие обеспечивают разложение большей части целлюлозы. Частицы древесины поглощаются ими, и целлюлоза переваривается внутриклеточно с выделением углекислого газа .и водорода. Основными продуктами расщепления (как и в рубце жвачных) являются летучие жирные кислоты, в первую очередь уксусная, которая всасывается в задней кишке. Без простейших термиты переваривать древесину не могут.
Бактерии играют менее заметную роль в кишечнике термитов, чем в рубце жвачных, но они, по-видимому, участвуют в двух особых случаях мутуализма.
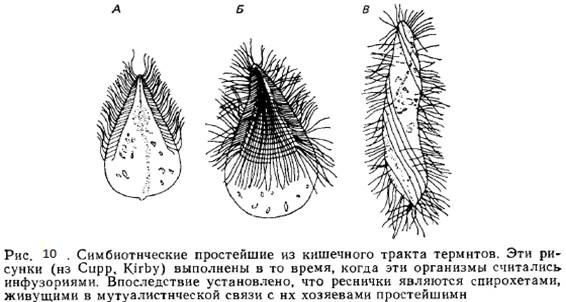
1. Важными представителями бактериальной флоры являются спирохеты; они вместе с палочковидными формами концентрируются на поверхности жгутиконосцев. В кишечнике одного из термитов (Mastoterma paradoxa) обнаружены спирохеты, которые, двигаясь синхронно, фактически продвигали вперед простейшее. Они настолько хорошо заметны, что когда-то таких жгутиконосцев считали инфузориями (рис. 10). Это взаимодействие бактерий и простейших мутуалистическое — спирохеты получают от жгутиковых .питательные вещества ,и одновременно придают им подвижность. Итак, в данном случае пара видов мутуалистически обитает в третьем виде.
2. Бактерии в кишечнике термитов способны фиксировать газообразный азот (N2
). Кливленд предполагал такую возможность еще в 1925 г., но не мог получить доказательств. Впоследствии Брезнак (Breznak, 1975) показал, что фиксация азота действительно происходит в кишечнике термитов и прекращается, если термита накормить антибактериальным препаратом. Когда в пище увеличивается содержание азота, скорость его фиксации также резко падает.
Существует множество тщательно выполненных исследований, на результате которых установлено существование симбионтов (главным образом, бактерий) внутри клеток животных (особенно насекомых). Например, огромные скопления бактерий неизменно присутствуют в жировых телах тараканов. Эти бактерии передаются через самку, скапливаясь сначала вокруг ооцитов, а затем проникая в цитоплазму яйца. Таким образом, этот симбиоз наследственный, передающийся через материнский организм. Сходным образом, в специализированных образованиях тлей (мицетомах) присутствуют дрожжеподобные симбионты, которые передаются потомству через яйца. Культивировать таких симбионтов в большинстве случаев невозможно, и их обмен веществ не изучен. Однако, иногда удавалось убивать их антибиотиками, после чего у хозяина наблюдались симптомы нарушения ряда метаболических функций, которые, по-видимому, должны обеспечиваться симбионтом. Так, кровососущий клоп Rhodnius prollxus в их отсутствие страдает от недостатка
витамина В, а у таракана-прусака (Blattella gertnanica) замедляется развитие, но при добавлении в пищу определенных полипептидов может быть восстановлена нормальная скорость роста. Предполагается, что эти вещества вырабатываются симбионтом, т. е. и эта взаимосвязь мутуалистическая.
То, что у большинства высших растений называют корнями, на самом деле микориза («грибокорень») — тесный мутуализм грибов и корневой ткани, при котором грибы помогают хозяевам получать минеральное питание, а сами берут у растения часть необходимого им органического углерода ( Harley, Smith, 1983). Лишь представители очень немногих
семейств, например, крестоцветные, не образуют такой ассоциации. У большинства мхов, папоротников, плаунов, голосеменных и покрытосеменных обнаружены ткани, более или менее тесню
переплетенные с мицелием грибов. Все доминанты основных типов растительности на Земле — лесные деревья, травы и кустарники — имеют хорошо выраженную микоризу. Ископаемые остатки древнейших наземных растений позволяют предполагать их тесную связь с грибами. У этих форм еще нет корневых волосков, а в некоторых случаях — даже корней, поэтому заселение суши могло зависеть от присутствия грибов-мутуалистов.
Существуют различные формы микоризы, мы рассмотрим эктомикоризу.
3.6.1. Эктомикориза
Эктомикоризные грибы нуждаются о растворимых углеводах.
Такая «покровная» микориза чаще всего встречается на корнях деревьев и образуется грибами из групп базидиомицетов или аскомицетов, многие из которых можно выращивать и в отсутствие хозяина. Заселенные ими корни обычно сконцентрированы в верхнем слое почвы, мицелий разрастается в подстилке и образует крупные выступающие над поверхностью плодовые тела, рассеивающие огромное количество переносимых ветром спор. Гриб покрывает корень своей «тканью» и обычно вызывает в нем морфогенетические изменения. Заселенные корни прекращают апикальный рост, остаются укороченными и часто дихотомически ветвятся, тогда как мицелий разрастается между клетками их коры, обеспечивая тесный контакт гриба и хозяина. Корни, проникающие глубже в почву, в слои, менее богатые органическим веществом, остаются незаселенными и продолжают нормально апикально удлиняться.
Грибам, обволакивающим корни растения-хозяина, в качестве источника углерода необходимы растворимые углеводы, и в этом отношении они отличаются от большинства своих свободноживущих, т. е. несимбиотических родичей, расщепляющих целлюлозу. Микоризные грибы по крайней мере часть своих потребностей в углероде удовлетворяют за счет хозяев. Мицелий всасывает из почвы минеральные биогены, и в настоящее время нет сомнений в том, что он активно снабжает ими растение- хозяина. В исследованиях с использованием радиоактивных меток обнаружено, что фосфор, азот и кальций по гифам грибов могут попадать в корни, а затем в побеги. Удивительно, что микориза, по-видимому, не менее эффективно действует и без гиф, отходящих от окутывающей корень «оболочки» из мицелия. Следовательно, сама эта «оболочка» должна обладать хорошо развитыми способностями поглощать питательные вещества и передавать их растению.
4. Эволюция субклеточных структур при симбиозе
Мутуализм — возможный источник эволюции эукариот.
Мы видели, что как среди растений, так и среди животных существуют весьма разнообразные взаимоотношения, которые можно считать мутуалистическим симбиозом. Сюда относятся ассоциации двух совершенно различных организмов, связанных поведенческими реакциями, но проводящих часть своего жизненного цикла независимо друг от друга и сохраняющих индивидуальные особенности (бычки и креветки, бабочка голубянка и муравьи). Далее по уровню сложности следуют экосистемы
типа хемостата (строго внешние по отношению к тканям) в рубце жвачных и слепой кишке термитов; затем — межклеточная эктомикориза . Эти стадии можно расценивать как последовательные этапы интеграции - сначала отдельных членов сообщества, а затем как бы частей одного «организма». Нетрудно представить, что некоторые из существ, считающихся едиными организмами, возникли в результате симбиоза приведшего к безвозвратной утере индивидуальности партнеров.
На этой точке зрения особенно настаивает Маргулис (Margulis 1975). Она считает, что переломные этапы в эволюции основных групп высших растений и животных сопровождались включением в состав их клеток прокариот (так называемая «последовательно-эндосимбиотическая теория эволюции эукариотической клетки»). «Первый шаг на пути возникновения эукариот от прокариот был сделан, когда внутрь клетки ферментативного анаэроба внедрилась эубактерия, в которой осуществлялся цикл Кребса (промитохондрия),... в результате чего образовались содержащие митохондрии амебоиды, давшие начало всем остальным эукариотам. Те в свою очередь приобрели поверхностные подвижные бактерии, превратившиеся в жгутики и реснички. Позднее поглощение клеток сине-зеленых водорослей могло бы привести к появлению автотрофного «организма», предка всего царства растений. Эта последовательность событий является чисто гипотетической и вызывает много скептических замечаний, но, по крайней мере отчасти, она может соответствовать действительности. До сих пор существуют близкие к предполагаемым симбионтам формы, включая бактерию Paracoccus denitrificans, рядом признаков напоминающую гипотетического свободноживущего предшественника митохондрии.
На первый взгляд создать математические модели мутуалистических взаимоотношений нетрудно. Для этого уравнение Лотки—Вольтерры, описывающее рост популяции одного вида, т. е. dN1
dt = r1
*N1
{K1
—N1
K1
), следует видоизменить, введя коэффициент (+α*N2
) /K1
, определяющий, наcколько особи вида 2 способствуют увеличению скорости роста популяции вида 1. Такое же уравнение может быть получено для вида 2 с использованием коэффициента(+β*N1
)/K2
,в этих уравнениях учитывается возможность увеличения численности каждого вида в присутствии другого вида, т. е. признак истинно мутуалистических отношений. Однако такая модель воспроизводит ситуацию, в которой численность обоих мутуалистов взрывообразно и неограниченно возрастает. Очевидно, что это нереалистично. Другие попытки моделирования мутуализма приводят к заключению, что такое взаимодействие довольно нестабильно. Но большая часть данных, полученных в естественных условиях, показывает его высокую устойчивость и способность быстро, восстанавливаться после нарушений. Почти наверняка неудача этих моделей объясняется их подходом к мутуализму как к зеркальному отражению конкуренции; в них предусматривается, что ограничения роста популяции, существующие для отдельного вида, при мутуализме ослабятся. В действительности же большинство случаев мутуализма, вероятно, не ослабляет ограничения, а устраняет, замещая другими. Например, если у кишечнополостного появляется фотосинтезирующий эндосимбионт, углеродное питание перестает быть лимитирующим рост фактором, и на первый план, по-видимому, выходят иные ограничения. Такое состояние может быть вполне устойчивым.
В биологии мутуалистов (особенно вступающих в тесные физические взаимосвязи) существует много характерных черт, отличающих их от большинства других организмов. По своей биологии они резко отличаются от паразитов и свободноживущих сородичей.
1.Жизненный цикл специализированных мутуалистов удивительно прост (в отличие прежде всего от большинства паразитов).
2.У эндосимбиотических мутуалистов половое размножение,
по-видимому, подавлено, особенно по сравнению с паразитами и свободноживущими сородичами.
3.У эндосимбионтов нет выраженной стадии расселения. Если оно и происходит, то часто у обоих партнеров вместе (например, когда молодая самка муравьев, покидая старую колонию, чтобы основать новую, берет с собой инокулят грибов, или когда гриб и водоросль у многих лишайников объединяются в расселительную структуру). Исключение из этого общего правила — споры из плодовых тел эктомикоризных грибов, но такие
грибы большую часть своей жизни могут проводить как свободноживущие организмы (или как паразиты). Это различие является особенно сильным между мутуалистамн и паразитами: популяционная динамика последних в большинстве случаев определяется расселением.
4.При мутуализме, по-видимому, не бывает ничего похожего на эпидемии, свойственные паразитизму. По сравнению с паразитами популяции мутуалистов представляются весьма стабильными.
5.В популяциях мутуалистов число эндосимбионтов приходящихся на одного хозяина, оказывается удивительно постоянным.
6.По-видимому, у организмов, образующих мутуалистический симбиоз, экологическая амплитуда (и ширина ниши) обычно (возможно, всегда) больше, чем у каждого из них при одиночном существовании. Этим мутуализм также отличается от паразитизма, когда экологическая амплитуда хозяина а присутствии паразитов обычно сужается.
7. Удивительно, что специализация по хозяевам у мутуалистов часто оказывается вполне гибкой — мутуализм муравьев и растений с нектарниками, водорослей и грибов в лишайниках, растений и опылителей и т. д. часто включает пары видов, которые могут быть мутуалистически связаны с несколькими, иногда многими другими видами. Строгая специализация не является здесь правилом.
Безусловно, экологи недооценивали значение мутуализма даже в большей степени, чем паразитизма. Их внимание было, вероятно, чрезмерно сосредоточено на экологии конкурентных отношений и взаимодействий типа хищник—жертва. Одна из причин этого, по-видимому, в том, что многие мутуалистическне отношения представляются чем-то из ряда вон выходящим, своего рода прихотью естественной истории, что всегда вызывает пренебрежение серьезных теоретиков. Мутуализм слишком хорошо подходит для занимательно-описательного подхода к объяснению сущности явлений, при котором любой объект рассматривается как деталь безупречно отлаженного механизма природы.
Многие работы по мутуализму представляют собой сборники
курьезных случаев, и данная глава о основном отражает такую ситуацию. Но изучение мутуализма затрагивает одну из наиболее фундаментальных проблем экологии. Являются ли сообщества организмов более или менее тесно коэволюционирующими образованиями? Возникают ли у них особые свойства в ходе эволюции взаимодействий? Многие примеры, по-видимому, подкрепляют точку зрения, согласно которой группы из двух или большего числа видов проявляют тенденцию к объединению во взаимовыгодные ассоциации, что приводит к образованию своего рода сверхорганизмов. Мы до сих пор почти ничего не знаем о том, как широко распространен такой
высокоразвитый тип мутуализма; возможно, в не столь выраженной форме он объединяет в природе функционирование целых сообществ. Являются ли эти примеры крайним выражением широко распространенного явления или только исключениями из правила, курьезами, уродствами? Одно из интересных предположений состоит в том, что в умеренных зонах, в отличие от тропиков, мутуализм редко бывает облигатным. Не (исключено, что роль факультативных мутуалистов в структуре сообществ или гильдий гораздо значительнее, чем предполагалось ранее.
На Земле организмы не могут существовать изолированно друг от друга. Между ними возникают различные типы взаимоотношений. Я рассмотрела мутуализм.
В начале представлены определения типов взаимоотношений, которые встречаются в природе, между живыми организмами. Затем рассмотрен, непосредственно, более подробно сам мутуализм и его типы, такие как сотрудничество и симбиотический мутуализм. Представлены их примеры, одни из немногих существующих в природе.
Так как считается, что мутуализм сыграл немалую роль в эволюции, то в моей работе представлена отдельная небольшая глава, рассматривающая эту тему.
В биологии мутуалисты проявляют схожие черты, которые отличают их от других большинства организмов. Последняя глава посвящена обобщению этих характерных черт.
Мутуализм важен для всего живого, поэтому он так распространен в природе и играет важную роль в поддержании разнообразия видов на Земле.
1. Бигон ,М. Экология. Особи популяции и сообщества[текст]:в 3-ех томах.Т.3/М.Бигон,Дж.Харпер,К.Таунсент;перевод с англ. В.Н.Михеева,М.А.Сметкова.-место М:Мир,1989.-667с.
2. Одум,Ю.Основы экологии[текст]/Ю.Одум;перевод с англ.Н.П.Наумнова,-место М:Мир,1975.-741с.
3. Коробкин ,В.И,Передельский Л.В.Экология: учебник для вузов/В.И.Коробкин,Л.В.Передельский.-Изд.12-е,доп. и перераб.-Ростов н/Д:Феникс,2007.-602,[1]с.-(Высшее образование)
4. Степановских А.С.Экология :учебник для вузов.-М.:Юнити-Дана.2001.-703с.
5. Чернова ,Н.М,Былова ,А.М.Общая экология.-М:Дрофа.2004.-416с.
|

