МИНИСТЕРСТВО ОБРАЗОВАНИЯ
РОССИЙСКОЙ ФЕДЕРАЦИИ
ТУЛЬСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ
Кафедра аэрологии, охраны труда и окружающей среды
Л.Н. Савинова
доцент, кандидат химических наук
Э К О Л О Г И Я
Конспект лекций
количество часов 34
Тула, 2000
СОДЕРЖАНИЕ
Лекция 1. Краткая история и предмет экологии.
Лекция 2.Экологическая система. Принципы и концепции.
Лекция 3. Энерг
ия в экологических системах.
Лекция 4.Энергия и продуктивность.
Лекция 5.Пищевые цепи, пищевые сети и трофические уровни.
Лекция 6. Трофическая структура и трофическая функция экосистемы.
Лекция 7. Биосфера как глобальная экосистема.
Лекция 8.Биогеохимические циклы
Лекция 9. Круговороты основных биогенных элементов:
глобальный круговорот воды и углерода.
Лекция 10. Круговороты основных биогенных элементов:
круговорот кислорода.
Лекция 11.Круговороты азота и серы .
Лекция 12.Осадочный цикл .
Лекция 13.Пути возвращения веществ в круговорот: коэффициентвозврата.
Лекция 14. Воздействие среды обитания на биоту.
Лекция 15. Абиотические факторы среды обитания
.
Лекция 16. Биотические отношения и роли видов в экосистеме.
Лекция 17.Развитие и эволюция экосистемы.
Лекция 1
Краткая
история и предмет экологии
1. Краткая история экологии.
2. Предмет экологии.
3. Подразделения экологии.
4. Иерархия уровней организации жизни.
5. Принцип эмерджентности.
Экология - этонаука о взаимоотношениях живых существ между
собой и с окружающей их неорганической природой,
о связях в надорганизменных
системах, о структуре и функцио
нирован
ии этих систем.
Экология как наука сформировалась лишь в середине прош
лого столетия, после того, как были накоплены сведения о многообразии
живых о
рганизмов
на Земле, об особенностях их образа жизни. Возникло понимание, что не только строение и развитие организмов, но и взаимоотношения их со средой обитания подчинены
определенным закономерностям, которые заслуживают специального и тщательного изучения.
Термин «экология» ввел известный немецкий зоолог Э. Геккель,
который в своих трудах «Всеобщая морфология организмов» (1866) и «Естественная история миротворения» (1868)
впервые попытался дать определение сущности новой науки. Слово «экология» происходит от греческого oikos, что означает «жилище», «местопребывание», «убежище». Э.Геккель
определял экологию как «общую науку об отношениях организмов к окружающей среде, куда мы относим в широком смысле все условия существования. Они частично органической, частично неорганической природы; но как те, так и другие имеют весьма большое значение
для форм организмов, так как они принуждают приспосабливаться к себе». По Геккелю,
экология представляет собой науку о «домашнем быте» живых организмов, она призвана исследовать «все те запутанные взаимоотношения, которые Дарвин условно обозначил как борьбу за существование».
Забиваем Сайты В ТОП КУВАЛДОЙ - Уникальные возможности от SeoHammer
Каждая ссылка анализируется по трем пакетам оценки: SEO, Трафик и SMM.
SeoHammer делает продвижение сайта прозрачным и простым занятием.
Ссылки, вечные ссылки, статьи, упоминания, пресс-релизы - используйте по максимуму потенциал SeoHammer для продвижения вашего сайта.
Что умеет делать SeoHammer
— Продвижение в один клик, интеллектуальный подбор запросов, покупка самых лучших ссылок с высокой степенью качества у лучших бирж ссылок.
— Регулярная проверка качества ссылок по более чем 100 показателям и ежедневный пересчет показателей качества проекта.
— Все известные форматы ссылок: арендные ссылки, вечные ссылки, публикации (упоминания, мнения, отзывы, статьи, пресс-релизы).
— SeoHammer покажет, где рост или падение, а также запросы, на которые нужно обратить внимание.
SeoHammer еще предоставляет технологию Буст, она ускоряет продвижение в десятки раз,
а первые результаты появляются уже в течение первых 7 дней.
Зарегистрироваться и Начать продвижение
Как и большинство наук, экология имеет длительную предысторию. Ее обособление представляет собой естественный этап роста
научных знаний о природе. Выделившись в системе других естественных наук, экология и сейчас продолжает развиваться, обогащая свое содержание и расширяя задачи. Современная экология является теоретической основой рационального природопользования, ей принадлежит ведущая роль в разработке стратегии
взаимоотношений природы и человеческого общества.
Накопление сведений об образе жизни, зависимости от внешних условий, характере распределения животных и растений началось очень
давно. Первые попытки обобщения этих сведений мы встречаем в трудах античных философов. Аристотель (384-322 до н. э.)
описал свыше 500 видов известных ему животных и рассказал об их поведении, например о миграциях и зимней спячке рыб, перелетах птиц, строительной деятельности жи
вотных, паразитизме кукушки, способе самозащиты у каракатицы и т. д.
Ученик Аристотеля, «отец ботаники» Теофраст
Эрезийский
(371-280 до н. э.)
привел сведения о своеобразии растений
в разных условиях, зависимости их формы и особенностей
роста от почвы и климата.
В средние века интерес к изучению природы ослабевает, заменяясь
господством богословия и схоластики.
Великие географические открытия в эпоху Возрождения, колонизация новых стран послужили толчком к развитию систематики. Описание растений и животных, их внешнего и внутреннего строения, разнообразия форм - главное содержание биологичес
кой науки на ранних этапах ее развития. Первые систематики - А. Цезальпин
(1519-1603), Д.
Рей (1623-1705), Ж.
Турнефор
(1656-1708) и другие сообщали и о зависимости растений от условий произрастания или возделывания, о местах их обитания и т. п. Сведения о поведении, повадках, образе жизни животных, которыми сопровождалось описание их строения, называ
ли «историей» жизни животных.
Сервис онлайн-записи на собственном Telegram-боте
Попробуйте сервис онлайн-записи VisitTime на основе вашего собственного Telegram-бота:
— Разгрузит мастера, специалиста или компанию;
— Позволит гибко управлять расписанием и загрузкой;
— Разошлет оповещения о новых услугах или акциях;
— Позволит принять оплату на карту/кошелек/счет;
— Позволит записываться на групповые и персональные посещения;
— Поможет получить от клиента отзывы о визите к вам;
— Включает в себя сервис чаевых.
Для новых пользователей первый месяц бесплатно.
Зарегистрироваться в сервисе
В ХУ
II-ХУII
I
вв.экологические сведения составляли нередко значительную часть в работах, посвященных отдельным группам живых организмов, например в трудах А.Реомюра о насекомых (1734),
Л. Трамбле
о гидрах и мшанках (1744), или в описаниях
путешествий, предпринимаемых натуралистами.
Много путешествий по неизведанным краям России было организовано
в XVIII
в. В трудах С.Л. Крашенинникова,
И.И. Лепех
ин
а, П.
С. Палласа и других русских географов, и
натуралистов указывалось на взаимосвязанные изменения кли
мата, растительности и животного мира в различных частях нашей обширной страны. П.
С. Паллас в своем капитальном труде «Зоография» подробно описал образ жизни 151 вида млекопи
тающих и 425 видов птиц и такие биологические явления, как ми
грации, спячка, взаимоотношения родственных видов и т. п.
Проблема влияния внешних условий на строение животных была поставлена во второй половине XVIII в. в трудах французского естествоиспытателя Ж. Бюффона
(1707-1788). Считая возможным «перерождение» видов, Бюффон
полагал основными причинами превращения одного вида в другой влияние таких внеш
них факторов, как «температура климата, качество пищи и гнет одомашнивания».
Жан Батист Ламарк
(1744-1829), автор первого эволюционного
учения, считал, что влияние «внешних обстоятельств» - одна из самых важных причин приспособительных
изменений организмов, эволюции животных и растений. Дальнейшему развитию экологического мышления способствовало появление в начале XIX столетия биогеографии. Труды
Александра Гумбольдта (1807) определили новое экологическое направление в географии растений. А. Гумбольдт ввел в науку представление о том, что «физиономия» ландшафта определяется внешним обликом растительности. В сходных зональных и вертикально-поясных географических условиях у растений разных таксономических
групп вырабатываются сходные «физиономические»
формы, т. е.
одинаковый внешний облик; по распределению и соотношению этих форм можно судить о специфике физико-географической среды. Появились первые специальные работы, посвященные влиянию климатических факторов на распространение и биологию животных, например книга немецкого зоолога К. Глогера
об изменениях птиц под влиянием климата (1833) и датчанина Т. Фабера
об особенностях биологии северных птиц (1826), К. Бергмана
о географических закономерностях в изменении размеров теплокровных животных (1848). А. Декандоль
в «Географии растений» (1855) подробно описан влияние отдельных факторов среды (температуры, влажности, света, типа почвы, экспозиции склона) на растения и обратил внимание на повышенную экологическую пластичность растений но
сравнению с животными.
Профессор Московского университета К.Ф. Рулье
(1814- 1858) широко пропагандировал необходимость развития особого направления в зоологии, посвященного всестороннему изучению и объяснению жизни животных, их сложных взаимоотношений с окружающим миром. К.Ф. Рулье
подчеркивал, что в зоологии наряду с классификацией отдельных органов нужно производить «
разбор явлений образа жизни». При этом следует различать явления жизни особи (выбор и запасание
пищи, выбор и постройка жилища и т. п.)
и «явления жизни общей» (взаимоотношения родителей и потомства, законы количественного размножения животных, отношение к животным того же вида и других видов, «жизнь в одиночестве», «жизнь в товариществе», «жизнь в обществе», отношения животных к растениям, почве, к физическим условиям среды). Наряду с этим следует специально изучать периодические явления в жизни животных - линьку, спячку, сезонные перемещения и т. д.
Таким образом, К.Ф. Рулье
разработал широкую систему экологического исследования животных, «зообиологии»,
в его понимании, и оставил ряд трудов типично экологического содержания, например типизацию общих особенностей водных, наземных и роющих позвоночных и т. д.
Взгляды К.Ф. Рулье
глубоко повлияли на направление и характер работ его учеников, одним из которых был Н.
А. Северцов (1827-1885). Его труд «Периодические явления в жизни зверей, птиц и гад Воронежской губернии» (1855) был первым в России глубоким экологическим исследованием животного мира отдельного региона.
В
1859 г. появилась книга Ч. Дарвина «Происхождение видов путем естественного отбора, или Сохранение благоприятствуемых
пород в борьбе за жизнь». Ч. Дарвин показал, что «борьба
за существование» в природе, под которой он понимал все формы противоречивых связок вида со средой, приводит к естественному
отбору, т. е.
является движущим фактором эволюции. Стало ясно, что взаимоотношения живых существ и связи их с неорганическими компонентами среды («борьба за существование») - большая самостоятельная область исследований.
Термин «экология» прижился не сразу и получил всеобщее признание лишь к концу XIX в. Во второй половине XIX столетия содержанием экологии было в основном изучение образа жизни животных и растений и их адаптации к климатическим условиям: температуре и световому режиму, влажности ит. д.
В этой области был сделан ряд важных обобщений. Продолжая «физиономическое»
направление А. Гумбольдта, датский ботаник Е.
Вар
минг
в книге «Ойкологическая
география растений» (1895) обосновал понятие о жизненной форме растения. А.Н. Бекетов
(1825-1902) выявил связь особенностей анатомического и морфологического строения растений с их географическим распространением и указал на значение физиологических исследований в экологии. А. Ф. Миддендорф,
изучая общие черты строения и жизни арктических животных, положил начало применению учения Гумбольдта к зоологическим объектам. Д.
Аллен (1877) нашел ряд общих закономерностей в изменении пропорций тела и
его выступающих частей и
в окраске североамериканских млекопитающих и птиц в связи с географическими изменениями климата.
Параллельно с этими исследованиями в конце 70-х
годов в экологии возникло новое направление. В 1877 г. немецкий гидробиолог К. Мебиус на основе изучения устричных банок Северного моря обосновал представление о биоценозе как о глубоко закономерном
сочетании организмов в определенных условиях среды. По Мебиусу, биоценозы, или природные сообщества, обусловлены длительной историей приспособления видов друг к другу и к сходной экологической обстановке. Изучение сообществ вскоре обогатилось методами учета количественных соотношений организмов. Учение о растительных сообществах обособилось в отдельную область ботанической экологии. Большую роль в этом сыграли труды русских ученых С.И. Коржинского
и И.К. Пачоского,
назвавшего новую науку «фитосоциологией».
Позднее она была переименована в «фитоценологию», а затем названа геоботаникой. Основные положения этой науки были разработаны в трудах Г.Ф. Морозова
и В.Н. Сукачева
на основе учения о лесе.
В начале XX столетия оформились экологические школы гидробиологов, фитоценологов,
ботаников и зоологов, в каждой из которых развивались определенные стороны экологической науки. На III ботаническом конгрессе в Брюсселе в 1910 г. экология растений официально разделилась на экологию особей (аутэкологию)
и экологию сообществ (синэкологию).
Это деление распространилось также на экологию животных, равно как и на общую экологию. Появились первые экологические сводки - руководства
к изучению экологии животных Ч. Адамса
(1913), книги В. Шелфорда
о сообществах наземных животных (1913), С.А. Зернова по гидробиологии (1913). В 1913-1920 гг. были организованы
экологические научные общества, основаны журналы, экологию начали преподавать в университетах.
К 30-м
годам, после разносторонних исследований и дискуссий, выкристаллизовались
основные теоретические представления в области биоценологии: о границах и структуре биоценозов, степени устойчивости, возможности саморегуляции
этих систем. Углубля
лись исследования типов взаимосвязей организмов, лежащих в ос
нове существования
биоценозов. Разрабатывалась соответствующая терминология.
Для развития идей общей биоценологии большое значение имели в нашей стране фитоценологические
исследования В.Н.
Сукаче
ва, Б.А. Ке
ллер
а, В.В. Алех
ин
а, Л.Г. Р
амен
ског
о, А.Л. Шенн
икова, за рубежом - Ф.
Клемен
тса в Америке, К. Раункиера
в Дании, Г. Дю
Рие в Швеции, И. Браун - Бланке
в Швейцарии. Были созданы разнообразные системы классификации растительности на основе морфологических (физиономических),
эколого-морфологических, динамических и других особенностей сообществ, разработаны представления об экологических индикаторах, изучены структура, продуктивность, динамические связи фитоценозов.
В разработку физиологических основ экологии растений, продолжая традиции К.А. Тимирязева, много ценного вн
ес Н.
А. М
аксим
ов.
В 30-40-х
годах появились новые сводки по экологии животных
с изложением теоретических проблем общей экологии: К. Фридерикса
(1930), Ф. Боденгеймера (1938)
и др. Большая роль в развитии общей экологии принадлежит Д.Н. Кашкарову,
опубликовавшему в
1933 г. сводку «Среда и с
ообщество»,
а позднее (1938) создавшему первый в нашей стране учебник по
основам экологии животных. Разрабатывались биоценологические
основы паразитологии ( В.В. Д
оге
ль, Е.Н. П
авловский,
В.Н. Беклемишев).
В России для экологии
характерна практическая направленность, глубокая разработка прикладных аспектов.
В 30-х
годах оформилась новая
область экологической науки - популяционная
экология. Основоположником ее следует считать английского
ученого Ч. Элтона. В своей книге «Экология животных» (1927
) Элтон переключает внимание с отдельного орга
низма
на популяцию
как единицу, которую следует изучать самостоятельно,
так как на этом уровне выявляются свои особенности экологических
адаптаций и регуляций. Центральными проблемами популяционной
экологии стали проблемы внутривидовой организации
и дина
мики
численности.
Представления о популяциях стали особенно энергично развиваться в экологии после того, как оформилась популяционная
генетика, а в систематике стали рассматривать вид как сложную систему. Развитию популяционных
исследований сильно способствовали также запросы практики - острая необходимость разработки основ борьбы с вредителями и конкурентами в сельском и лесном хозяйстве, истощение запасов ряда ценных промысловых животных, открытие роли некоторых диких животных в распространении паразитов и возбудителей болезней человека и домашнего скота.
В развитие популяционной
экологии в нашей стране большой вклад внесли С.А. Северцов,
С.С. Шварц, Н.
П. Наумов,
Г.А. Викторов, работы которых во многом определяют современное состояние этой области науки.
Начало исследований популяций у растений было положено трудами Е.Н. Синской
(1948), много сделавшей по выяснению экологического и географического полиморфизма видов. Ряд вопросов популяционной
экологии растений разрабатывается в трудах Т.А. Работнова
и А.А. Уранова и их последователей.
Изучение популяционных
закономерностей по-новому помогло осознать роль видов в биоценозах, структурную организацию сообществ. Возникла плодотворная концепция «экологических ниш», тесно связывающая экологические и эволюционные вопросы. В ее разработке важная заслуга принадлежит западным ученым Дж. Гр
инне
ллу, Ч.Т. Э
лтон
у, Р. М
акар
тур
у, Д.
Xатчин
сон
у и советскому исследователю Г.Ф. Г
аузе.
Параллельно развиваются и другие области экологии, тесно связывающие эту науку с традиционными областями биологии. В развитие морфологической и эволюционной экологии животных большой вклад внес М.
С. Гиляров,
выдвинувший предположение, что почва послужила переходной средой в завоевании членистоногими суши (1949). Проблемы эволюционной экологии позвоночных животных нашли отражение в трудах С.С. Шварца.
И.С. Серебряковым
была создана новая, более глубокая классификация жизненных форм цветковых растений. Возникла палеоэкология,
задача которой - восстановление картины образа жизни вымерших форм.
С начала 40-х
годов в экологии возник принципиально новый подход к исследованию природных экосистем. В 1935 г. английский ученый А. Тенсли
выдвинул понятие экосистемы, а в 1942г.
В. Н. Сукачев
обосновал представление о биогеоценозе.
В этих понятиях нашла отра
жение
идея о единстве совокупности организмов с абиотическим окружением, о закономерностях, которые
лежат в основе связи всего сообщества и окружающей неорганической среды, о
круговороте вещества и превращениях энергии. Начались работы по точному учету продуктивности водных сообществ (Г.Г. Винберг,
1936). В 1942 г. американский ученый Р. Линдеман
опубликовал статью с изложением основных методов расчета энергетического баланса экологических систем. С этого периода стали принципиально возможными расчеты и прогнозирование
предельной продуктивности биоценозов в конкретных
условиях среды.
С развитием экосистемной
и популяционной
экологии более отчетливо стала вырисовываться специфика методов современной экологической науки. Основной инструмент экологического поиска представляют методы количественного анализа. Надорганизменные
объединения (популяции, сообщества, экосистемы) управляются
преимущественно количественными соотношениями особей, видов, энергетических потоков. Количественные изменения в струк
туре популяций и экосистем могут в корне переменить способы их функционирования, результаты деятельности. Развитие количест
венных методов исследования превращает экологию в точную науку, дает основы для математического моделирования, делает возможным научный прогноз. Это особенно важно для оценки устойчивости и продуктивности популяций и экосистем. В разработке теоретических основ биологической продуктивности
, начиная с 50-х
годов, принимают участие многие экологи, из которых особенно велики заслуги Г. Одума
и Ю. Одума, Р. Уитеккера, Р. Маргалефа
и других ученых. В нашей стране это направление наиболее успешно развивается в трудах гидробиологов и геоботаников.
Развитие экосистемного
анализа привело к возрождению на новой экологической основе учения о биосфере, принадлежащего крупнейшему естествоиспытателю XX в. В.И. Вернадскому, который в своих идеях намного опередил современную ему науку. Биосфера предстала как глобальная экосистема, стабильность и функционирование которой основаны на экологических законах обеспечения баланса вещества и энергии. Такой подход позволил ученым разных стран, работавшим с 1964 г. по общей Международной биологической программе (МБП), подсчитать максимальную биологическую продуктивность всей нашей планеты, т.е. тот природный фонд, которым располагает
человечество, и максимально возможные нормы изъятия продукции
для нужд растущего населения Земли.
Основным практическим результатом развития экосистемной
экологии явилось ясное осознание, сколь велика зависимость человеческого общества от состояния природы на нашей планете, необходимости
перестраивать экономику в соответствии с экологическими законами. Движение, которое лучше всего назвать как «всеобщая озабоченность проблемами окружающей среды», внезапно развернулось в течение двух лет, с 1968
по 1970 г. Казалось, все вдруг заинтересовались загрязнением среды, окружающей природой, ростом народонаселения, вопросами потребления пищи и энергии. Об этом свидетельствовали подробные материалы по проблемам окружающей среды в широкой прессе. Рост общественного интереса оказал глубокое влияние на академическую экологию. До 1970 г. на
экологию смотрели главным образом лишь как на одно из подразделений биологии. Экологи входили в штат биологических факультетов, курсы экологии, как правило, имелись только в программе подготовки биологов. Хотя и сейчас экология уходит своими корнями в биологию, она уже вышла из ее рамок, оформившись в принципиально новую интегрированную дисциплину, связывающую физические и биологические явления и образующую мост между естественными и общественными науками.
На современном этапе
развития экология обязана изучать не только связи организмов и законы функционирования надорганизменных
систем, но и обосновывать рациональные формы взаимоотношений
природы и человеческого общества. Растет социальная роль экологических знаний. Основные цели развития фундаментальных исследований в области экологии определяются острыми народнохозяйственными проблемами: необходимостью интенсифицировать производство и повышать экономическую эффективность использования природных ресурсов, сохраняя при этом состояние окружающей среды. На первый план выдвигаются вопросы биологической продуктивности и стабильности природных и искусственных сообществ. Эти проблемы могут быть
решены только совместными усилиями экологов всех стран, поэтому широко реализуется международное сотрудничество в области глобальной экологии. В настоящее время ясно осознана опасность экологического кризиса, возможности катастрофических неравновесных преобразований планетарной системы в связи с широкой экстенсивной хозяйственной деятельностью человека. Возможности предотвращения этого кризиса могут быть найдены
только на основе развития экологических знаний. Действенная сила экологических знаний помогает правильно эксплуатировать природные ресурсы, управлять численностью популяций, находить новые решения сельскохозяйственных проблем, новые принципы
организации промышленных производств.
Современная экология представляет собой разветвленную систему наук. Она делится на общую экологию, изучающую закономерности связи со средой, присущие всем группам организмов, и на
частные направления, по экологической специфике отдельных групп (экология микроорганизмов, растений, млекопитающих, птиц, рыб, насекомых и т. п.).
В ней выделяются такие области, как аут- и синэкология,
экология популяций. Физиологическая экология выявляет закономерности физиологических изменений, лежащих в основе адаптации организмов. В последние годы развивается биохимическая экология, внимание которой направлено
на молекулярные механизмы приспособительных преобразований
в организмах в ответ на изменение среды. Палеоэкология
изучает экологические связи вымерших групп, эволюционная экология - экологические механизмы преобразования популяций, морфологическая экология - закономерности строения органов и структур в зависимости от условий обитания. Основной предмет геоботаники - закономерности сложения и распределения фитоце
нозов. Экологической наукой является гидробиология. Выделяют также экологию наземных экосистем, экологию ландшафтов и т. д. Особую область составляет математическая экология, задачей ее является перевод эмпирически накопленных сведений и закономерностей в математические модели, которые позволяют прогнозировать
состояние и поведение популяций и сообществ. В последнее время развивается экология человека, включающая в себя и целый ряд социальных проблем.
Иерархия уровней организации
.
Вероятно, лучше всего можно определить содержание современной экологии, исходя из концепции уровней организации жизни, которые составляют своеобразный «биологический спектр», как это показано на рис. 1.
Биотические Гены Клетки Органы Организм Популяции Сообщества
компоненты ßßßßßß
 + +
Абиотические вещество ~~~ энергия
компоненты
= ßßßßßß
Биосистемы генети- клеточ- системы системы популяцион- экосистемы
ческие ные органов организ- ные системы
системы системы мов
Рис. 1.Спектр уровней организации.
Сообщество, популяция, организм, орган, клетка и ген - основные уровни организации жизни; на рис. 1. они
расположены в иерархическом порядке - от крупных систем к малым (иерархия
- это расположение ступенчатым рядом). На каждой ступени, или уровне, в результате взаимодействия с окружающей физической средой (энергией и веществом) возникают характерные функциональные
системы. Под системой мы будем подразумевать упорядочено
взаимодействующие и взаимозависимые компоненты, образующие единое целое.
Экология изучает главным образом те системы, которые расположены в правой части биологического спектра, т. е.
системы выше уровня организма. В экологии значение термина популяция,
первоначально обозначавшего группу людей, расширено и обозначает группы особей любого вида организмов. Точно так же сообщество(иногда называемое еще биотическим сообществом) в экологическом смысле включает все популяции, занимающие данный участок. Сообщество и неживая среда функционируют совместно, образуя экологическую систему, или экосистему.
Сообществу и экосистеме приблизительно соответствуют часто употребляемые в европейской и русской литературе термины биоценози биогеоценоз
. Биом
- удобный и широко используемый термин, обозначающий крупную региональную или субконтинентальную биосистему,
характеризующуюся каким-либо основным типом растительности или другой характерной особенностью ландшафта, например биом
лиственных лесов умеренного пояса. Самая крупная и наиболее близкая к идеалу в смысле «самообеспечения» биологическая система,
которую мы знаем, - это биосфера,или экосфера,
она включает все живые организмы Земли, находящиеся во взаимодействии с физической средой Земли как единое целое, чтобы поддерживать эту систему в состоянии устойчивого равновесия, получая поток энергии от Солнца.
Деление ступенчатого ряда, или иерархии, на компоненты во многих случаях искусственно, (например, системы “хозяин-паразит” или двухвидовая система взаимосвязанных организмов (сожительство гриба и водоросли, образующее лишайник) представляют собой промежуточные уровни между популяцией и сообществом) но иногда такое деление может быть основано на естественных разрывах. Так как каждый уровень в спектре биосистемы
«интегрирован», т. е.
взаимосвязан с другими уровнями, здесь нельзя найти резких границ или разрывов в функциональном смысле. Их нет даже между организмом и популяцией. Например, организм, изолированный от популяции, не в состоянии жить долго, точно так же, как изолированный орган не может длительное время сохраняться как самоподдерживающаяся
единица без своего организма. Подобным же образом сообщество не может существовать, если в нем не происходит круговорот веществ и в него не поступает энергия. Тот же аргумент можно привлечь для опровержения неверного представления о том, будто бы человеческая цивилизация
может существовать независимо от мира природы.
Принцип эмерджентности.
Важное следствие иерархической организации состоит в том, что по мере объединения компонентов, или подмножеств, в более «крупные функциональные единицы, у этих новых единиц возникают новые свойства, отсутствовавшие на предыдущем уровне. Такие качественно новые,
эмерджентные,
свойства экологического уровня или экологической единицы нельзя предсказать, исходя из свойств компонентов,
составляющих этот уровень или единицу. Иными словами, свойства целого невозможно свести к сумме свойств его частей. Хотя данные, полученные при изучении какого-либо уровня, помогают при изучении следующего, с их помощью никогда нельзя полностью объяснить явления, происходящие на этом следующем уровне; он должен быть изучен непосредственно.
Для иллюстрации принципа эмерджентности
приведем два примера, один из химии, другой из экологии. Водород и кислород, соединяясь в определенном соотношении, образуют воду, жидкость, совершенно непохожую по своим свойствам на исходные газы. А определенные водоросли и кишечнополостные
животные, эволюционируя совместно, образуют систему кораллового рифа, возникает эффективный механизм круговорота элементов питания, позволяющий такой комбинированной системе поддерживать высокую продуктивность в водах с очень низким содержанием этих элементов. Следовательно, фантастическая продуктивность и разнообразие коралловых рифов - эмерджентные
свойства, характерные только для уровня рифового сообщества.
При каждом объединении подмножеств в новое множество возникает по меньшей мере одно новое свойство; предлагается различать эмерджентные
свойства,
определение которых дано выше, и совокупные свойства,
представляющие собой сумму свойств компонентов. И те и другие - свойства целого, но совокупные свойства не включают новых или уникальных особенностей, возникающих при функционировании системы как целого. Рождаемость - пример совокупного свойства, поскольку она представляет собой лишь сумму индивидуальных рождений за определенный период, выраженную в виде доли или процента общего числа особей в популяции. Эмерджентные
свойства возникают в результате взаимодействия компонентов, а не в результате изменения природы этих компонентов. Части не «сплавляются», а интегрируются, обусловливая появление уникальных новых свойств.
Некоторые признаки, естественно, становятся более сложным
и и изменчивыми, когда по иерархии уровней организации (рис. 1.) продвигаешься слева направо, другие же,
напротив, часто становятся менее сложными и менее изменчивыми. Поскольку на всех уровнях функционируют гомеостатические
механизмы, а именно корректирующие и уравновешивающие процессы, действующие и противодействующие силы, амплитуда колебаний имеет тенденцию уменьшаться, когда мы переходим к рассмотрению более мелких единиц, функционирующих внутри крупных. Статистически разброс значений целого меньше суммы
разброса частей. Например, интенсивность фотосинтеза лесного сообщества менее изменчива, чем интенсивность фотосинтеза у отдельных листьев или деревьев внутри сообщества; объясняется это тем, что если в одной части интенсивность фотосинтеза снижается, то в другой возможно его компенсаторное
усиление. Если учесть эмерджентные
свойства и усиление гомеостаза
на каждом уровне, то станет ясно, что для изучения целого не обязательно знать все его компоненты. Это важный момент, поскольку некоторые исследователи считают, что не имеет смысла пытаться изучать сложные
популяции и сообщества, не изучив досконально составляющие его более мелкие единицы. Напротив, изучение можно начать с любой точки спектра при условии, что учитывается не только изучаемый,
но и соседние уровни, поскольку, как уже было сказано, некоторые свойства целого можно предсказать, исходя из свойств его частей (совокупные свойства), другие же нельзя (эмерджентные
свойства). Идеальное изучение какого-либо уровня системы включает изучение трехчленной иерархии: системы, подсистемы (соседний низший
уровень) и надсистемы
(следующий верхний уровень).
В соответствии со сказанным мы будем обсуждать принципы экологии на уровне экосистемы, уделяя достаточно внимания таким под системам , как популяция и сообщество, и такой надсистеме, как биосфера.
Лекция 2.
Экологическая система. Принципы и концепции.
1. Концепция экосистемы.
2. Изучение экосистем.
3. Стабильность экосистем.
Если мы хотим, чтобы наше общество перешло к целостному решению проблем, возникающих на уровне биомов и биосферы, то должны прежде всего изучать экосистемный уровень организации.
Концепция экосистемы
Живые организмы и их неживое (абиотическое) окружение неразделимо связаны друг с другом и находятся в постоянном взаимодействии. Любая биосистема, включающая все совместно функционирующие организмы (биотическое сообщество) на данном участке и взаимодействующая с физической средой таким образом, что поток энергии создает четко определенные биотические структуры и круговорот веществ между живой в неживой частями, представляет собой экологическую систему или экосистему.
Долговременное функционирование экосистемы обеспечивают три основных компонента - сообщество, поток энергии и круговорот веществ.
Поток энергии направлен в одну сторону; часть поступающей солнечной энергии преобразуется сообществом и переходит на качественно более высокую ступень, трансформируясь в органическое вещество, представляющее собой более концентрированную форму энергии, чем солнечный свет, но большая часть энергии деградирует, проходит через систему и покидает ее в виде низкокачественной тепловой энергии (тепловой сток). Энергия может накапливаться, затем снова высвобождаться или экспортироваться но ее нельзя использовать вторично.
В отличие от энергии элементы питания, в том числе биогенные элементы, необходимые для жизни (углерод, азот, фосфор и т. д.), и вода не только могут, но и должны использоваться многократно.
Все экосистемы, даже самая крупная - биосфера, являются открытыми системами: они должны получать и отдавать энергию. Разумеется, экосистемы, входящие в биосферу, также в разной степени открыты для потоков веществ, для иммиграции и эмиграции организмов. Поэтому концепция экосистемы должна учитывать существование связанных между собой и необходимых для функционирования и самоподдержания экосистемы среды на входеи среды на выходе: в концептуально законченную экосистему входит среда на входе, среда на выходе и система , т.е.
Экосистема = IE + S + OE.
Данная схема решает проблему, связанную с проведением границ рассматриваемой единицы, поскольку в этом случае не имеет значения, как мы вычленяем исследуемую часть экосистемы. Часто удобными оказываются естественные границы, например берег озера или опушка леса, или административные, например границы города, но эти границы могут быть и условными, если их точно определить геометрически. Конечно, экосистема не ограничена «ящиком» в центре схемы, поскольку если бы этот «ящик» был герметичным, то его живое содержимое (озеро или город) не вынесло бы такого заключения. Функционирующая реальная экосистема должна иметь вход и в большинстве случаев пути оттока переработанной энергии и веществ.
Изучение экосистем
При изучении больших сложных экосистем, таких, как озера и леса, экологи используют четыре основных подхода:
1) холистический(от греч. holos - целый) , который предполагает измерение поступлений и выхода энергии и различных веществ, оценку совокупных и эмерджентных свойств, а затем в случае необходимости - изучение его составных частей; экосистема рассматривается как черный ящик,т. е. как объект, функция которого может быть описана без выяснения его внутреннего содержания.
2) мерологический(от греч. meros - часть), при котором сначала изучаются свойства основных частей, а затем эти сведения экстраполируются на систему в целом. Очевидно, что важные эмерджентные свойства при мерологическом подходе могут быть упущены. Но, что самое главное, конкретный организм в разных системах может вести себя совершенно по-разному, и эта изменчивость, очевидно, связана с тем, как данный организм взаимодействует с другими компонентами экосистемы. Например, многие насекомые в агроэкосистеме являются опасными вредителями, а в своих естественных местообитаниях они не опасны, так как там их держат под контролем паразиты, конкуренты, хищники пли химические ингибиторы.
3) экспериментальные методы, т.е. нарушение тем или иным способом структуры или функции экосистемы в надежде, что реакция системы на такое нарушение позволит проверить гипотезы, основанные на наблюдениях. Экспериментальные методы - основа «стрессовой», или «пертурбационной» экологии.
4) методы моделирования. Модель-это абстрактное описание того или иного явления реального мира, позволяющее делать предсказания относительно этого явления. В своей простейшей форме модель может быть словесной или графической (неформализованной). Однако если мы хотим получить достаточно надежные количественные прогнозы, то модель должна быть статистической и строго математической (формализованной).
Моделирование обычно начинают с построения схемы, или графической модели, часто представляющей собой блок-схему. В работающей модели экологической ситуации имеется как минимум четыре ингредиента или компонента, а именно: 1) источник энергии или другая внешняя движущая сила,2) свойства, которые системоаналитики называют переменными состояний,3) направления потоков,связывающих «действа между собой и с действующими силами через потоки энергии и вещества; и 4)взаимодействия или функции взаимодействийтам, где взаимодействуют между собой силы и свойства, изменяя, усиливая или контролируя перемещения веществ и энергии или создавая эмерджентные свойства.
Характеристика хорошей модели должна включать три компонента: 1) анализируемое пространство (границы системы), 2) субсистемы (компоненты), считающиеся важными для общего функционирования, и 3) рассматриваемый временной интервал. После того как мы правильно определили экосистему, экологическую ситуацию или проблему и установили ее границы, мы выдвигаем доступную для проверки гипотезу или серию гипотез, которую можно принять или отвергнуть хотя бы предварительно, ожидая результатов дальнейших экспериментов или анализа. Более подробные сведения об экологическом моделировании можно найти в работах Холла и Дэя (1979), а также Медоуза (1982).
Стабильность экосистем
Стабильность экосистемы обеспечивается непрерывным потоком энергии, который задает и поддерживает круговороты веществ; а также развитыми информационными сетями, включающими потоки физических и химических сигналов, связывающих все части системы и управляющих (или регулирующих) ею как одним целым. В результате взаимодействия круговоротов веществ и потоков энергии, а также сигналов обратной связи от субсистем (когда часть сигналов с выхода поступает на вход) в экосистемах возникает саморегулирующийся гомеостаз без регуляции извне ( как это бывает в механических системах; например, в обычной системе регулировки температуры в помещении термостат управляет печью) или «постоянной точки» ( так у теплокровных животных регуляция температуры тела осуществляется специальным центром в мозгу). Управляющие функции экосистемы сосредоточены внутри нее и диффузны (а не направлены вовне и специализированы). В число управляющих механизмов, действующих на уровне экосистемы, входят микробные субсистемы, регулирующие накопление и высвобождение биогенных элементов, поведенческие механизмы и субсистемы «хищник-жертва», регулирующие плотность популяции, а также многие другие.
Помимо системы обратной связи стабильность обеспечивается избыточностью функциональных компонентов. Например, если в сообществе имеется несколько видов автотрофов, каждый из которых характеризуется своим температурным диапазоном функцнонирования, то скорость фотосинтеза сообщества в целом может оставаться неизменной, несмотря на колебания температуры.
По определению, стабильность - это свойство любого тела, заставляющее его возвращаться к исходному состоянию после того, как это тело выведено из состояния равновесия. Это представляется достаточно ясным, но на практике люди разных профессий (например, инженеры, экологи или экономисты) могут вкладывать в термин «стабильность» разный смысл, особенно при попытках оценить меру стабильности и выразить ее количественно. Для целей экологии можно выделить два «типа» стабильности.
Резистентная устойчивость- это способность экосистемы сопротивляться пертурбациям (нарушениям), поддерживая неизменной свою структуру и функцию. Упругая устойчивость - этоспособность системы восстанавливаться после того, как ее структура и функция были нарушены. Эти два типа стабильности связывает обратная зависимость - системе трудно одновременно развить оба типа устойчивости.
Так, калифорнийский лес из секвойи довольно устойчив к пожарам (для этих деревьев характерны толстая кора и другие адаптации), но если он все же сгорит, то восстанавливается очень медленно или вовсе не восстанавливается. Напротив, калифорнийские заросли чапарраля очень легко выгорают (низкая резистентная устойчивость), но быстро восстанавливаются за несколько лет (отличная упругая устойчивость). Как правило, при благоприятных физических условиях среды экосистемы в большей степени проявляют резистентную, а не упругую устойчивость, а в изменчивых физических условиях наблюдается прямо противоположное.
Гомеостатические механизмы функционируют в определенных пределах, за которыми уже ничем не ограничиваемые положительные обратные связи, усиливающие отклонения, приводят к гибели системы, если невозможно произвести дополнительную настройку. По мере нарастания стресса система, продолжая оставаться управляемой, может оказаться неспособной к возвращению на прежний уровень. На самом деле, согласно нашедшей широкое признание теории Холдинга (1973), для популяций и, как можно предположить, для экосистем характерно не одно, а несколько состояний равновесия и после стрессовых воздействий они часто возвращаются не в то состояние равновесия, из которого были выведены, а в другое. Вспомним, например, что значительное, хотя и не все количество СО2
, поступающей в атмосферу в результате деятельности человека, поглощается карбонатной и другими системами моря, но по мере увеличения притока СО2
в атмосфере устанавливаются новые равновесия на несколько более высоком уровне. В этом случае даже небольшое нарушение может иметь далеко идущие последствия. Во многих случаях подлинно надежный гомеостатический контроль устанавливается только после периода эволюционной «подгонки». Для новых экосистем (например, систем, создаваемых современным сельским хозяйством) или недавно сложившихся комплексов «паразит-хозяин» обычно характерны более резкие колебания и чрезмерный рост численности по сравнению со зрелыми системами, компоненты которых имели возможность приспособиться друг к другу.
Степень стабильности, достигаемая конкретной экосистемой, зависит не только от ее истории и эффективности ее внутренних управляющих механизмов, но и от характера среды на входе и, возможно, от сложности экосистемы. Как правило, экосистемы имеют тенденцию становиться сложнее в благоприятной физической среде, чем в среде со схоластическими (случайными, непредсказуемыми) нарушениями на входе, например штормами. Функциональная сложность, по-видимому, в большей степени, чем структурная, увеличивает стабильность системы, так как возрастает потенциально возможное число петель обратной связи; однако причинно-следственные взаимоотношения между сложностью и стабильностью изучены еще не достаточно.
Таким образом, одним из главных действующих лиц в экологии является поток энергии. Он обеспечивает стабильность экосистем; он - основа движения веществ в биогеохимических циклах. Поэтому мы свое изучение экологии начнем с поведения энергии в экосистемах.
Лекция 3.
Энергия в экологических системах.
1. Обзор фундаментальных концепций, связанных с энергией.
2. Жизнь как термодинамический процесс.
3. Энергетические характеристики среды.
Обзор фундаментальных концепций, связанных с энергией.
Важнейшим аспектом экологии
являются энергетические взаимоотношения
в экологических системах. Но прежде чем перейти к изучению энергетики экосистем, нам необходимо рассмотреть некоторые основы термодинамики.
Энергию определяют как способность производить работу. Свойства энергии описываются следующими законами.
Первый закон термодинамики, или закон сохранения энергии, гласит, что энергия может переходить из одной формы в другую, но она не исчезает и не создается заново. С точки зрения первого закона возможны и равновероятны любые процессы, в которых вместо исчезнувшего вида энергии появится эквивалентное количество другого вида. Так, первому закону не противоречило бы поднятие груза или закручивание какой-либо пружины за счет внутренней энергии окружающей среды. Почему, в самом деле, камень, лежащий на земле, не может подняться на какую-то высоту за счет охлаждения окружающего воздуха? Однако не поднимается. Переход теплоты от менее нагретого тела к более нагретому означал бы лишь перераспределение энергии внутри системы и также не противоречил первому закону. Однако известно, что сосуд с водой никогда не закипит на холодной плите. Иными словами, первый закон ничего не говорит о возможности и вероятности того или иного процесса, связанного с превращением энергии или ее перераспределением.
Между тем, если внимательно рассмотреть всевозможные процессы, протекающие в окружающем мире, а также проводимые нами самими, окажется, что их можно разбить на две существенно различающиеся группы. Во-первых, это процессы самопроизвольные, т.е. идущие сами собой. Для их проведения не только не затрачивается работа, но будучи поставленными в соответствующие условия, они сами могут произвести работу в количестве, пропорциональном происходящему изменению (например, переход теплоты от горячего тела к холодному, переход энергии заряженного аккумулятора в теплоту и т.д.). Самопроизвольные процессы ведут систему к состоянию равновесия, где силы, вызывающие процессы уравновешиваются. (например, выравниваются давление, температура, концентрация и т.д.). В случае попытки повернуть самопроизвольные процессы вспять, мы имеем дело уже с несамопроизвольными процессами. Они не идут сами собой. Для их проведения необходимо затратить работу в количестве, пропорциональном происходящему изменению. Критерии самопроизвольного или несамопроизвольного изменения системы, а также критерии равновесия устанавливает второй закон термодинамики.
Второй закон термодинамики, или закон энтропии, формулируется по-разному. Для наших целей более удобными являются следующие формулировки, которые мы приводим ниже:
процессы, связанные с превращениями энергии, могут происходить самопроизвольно только при условии, что энергия переходит из концентрированной формы в рассеянную (деградирует);
поскольку некоторая часть энергии всегда рассеивается в виде недоступной для использования тепловой энергии, эффективность самопроизвольного превращения кинетической энергии (например, света) в потенциальную (например, энергию химических соединений протоплазмы) всегда меньше 100 %.
Энтропия (от греческого entropia - поворот, превращение) - мера количества энергии, которая становится недоступной для использования, мера изменения упорядоченности, которая происходит при деградации энергии. Система обладает низкой энтропией, если способна создавать и поддерживать высокую степень внутренней упорядоченности за счет непрерывного рассеяния легко используемой, концентрированной энергии (например, света или пищи) и превращения ее в энергию, используемую с трудом (например, в тепловую).
Закон сохранения энергии и закон энтропии - это фундаментальные законы природы, имеющие универсальное значение. Насколько нам известно, из этих физических законов нет исключений и никакие технические изобретения не могут их нарушить. Любая искусственная или естественная система не подчиняющаяся этим законам обречена на гибель.
Легко показать, каким образом сформулированные фундаментальные физические концепции можно отнести к экологии. Все разнообразие проявлений жизни сопровождается превращениями энергии, хотя энергия при этом не создается и не уничтожается (первый закон термодинамики). Энергия, получаемая в виде света поверхностью Земли, уравновешивается энергией, излучаемой с поверхности Земли в форме невидимого теплового излучения. Сущность жизни состоит в непрерывной последовательности таких изменений, как рост, самовоспроизведение и синтез сложных химических соединений. Без переноса энергии, сопровождающего все эти изменения, не было бы ни жизни, ни экологических систем. Наша цивилизация - лишь одно из замечательных явлений природы, зависящих от постоянного притока концентрированной энергии светового излучения. Экология, по сути дела, изучает связь между светом и экологическими системами и способы превращения энергии внутри системы. Ибо отношения между растениями - продуцентами и животными - консументами, между хищниками и жертвой, не говоря уже о численности и видовом составе организмов в каждом местообитании, лимитируются и управляются потоком энергии, превращающейся из ее концентрированных форм в рассеянные.
Год за годом солнечное излучение распространяется в космическом пространстве. Какая-то часть этого излучения попадает на Землю, проходит через пленку атмосферы и становится доступной многим сотням экологических систем, покрывающим нашу планету и входящим в состав ее биосферы. Когда свет поглощается предметом, который при этом нагревается, световая энергия превращается в другую форму энергии - тепловую, то есть в энергию колебательных и поступательных движений молекул. В результате неравномерного поглощения солнечных лучей сушей и водой возникают теплые и холодные области - это служит причиной образования воздушных потоков, которые могут вращать ветряные двигатели и выполнять другую работу, скажем поднимать воду насосом против действия силы тяжести. Итак, в этом случае энергия света превращается в тепловую энергию земной поверхности, а затем в кинетическую энергию движущегося воздуха, которая выполняет работу подъема воды. При поднятии воды энергия не исчезает, а превращается в потенциальную, поскольку энергию, скрытую в поднятой воде можно снова превратить в какую-либо другую форму энергии, если дать воде опять упасть.
Энергия в какой-либо форме всегда пропорциональна количеству той формы энергии, в которую она переходит. "Потребленная энергия" не расходуется, она только переводится из состояния, в котором ее легко превратить в работу, в состояние с малой возможностью использования (бензин в баке автомобиля действительно расходуется, но энергия, содержащаяся в нем, не исчезает, а превращается в формы, уже неприродные для использования в автомобиле).
Так, попав на Землю, лучистая энергия солнца стремится превратиться в тепловую. Лишь очень небольшая часть световой энергии, поглощенной зелеными растениями, превращается в потенциальную энергию пищи, большая же ее часть превращается в тепло, покидающее затем и растение, и экосистему, и биосферу. Весь остальной живой мир получает необходимую потенциальную химическую энергию из органических веществ, созданных фотосинтезирующими растениями или хемосинтезирующими микроорганизмами. Например, животные поглощают химическую потенциальную энергию пищи и большую ее часть превращают в тепло, а меньшую вновь переводят в химическую потенциальную энергию заново синтезируемой протоплазмы. На каждом этапе передачи энергии от одного организма к другому значительная часть ее превращается в тепло, рассеивается в соответствии со вторым законом термодинамики - законом энтропии.
Мы проследили два пути превращения кинетической энергии солнечного света. Первый путь - превращения в энергию теплового излучения. И второй - путь поглощения солнечного света фотосинтезирующими организмами с продуцированием органического вещества. В каждом случае попытаемся понять, какую форму энергии, концентрированную или рассеянную, затрагивает превращение, на каких этапах превращение энергии происходит количественно, на каких нет:
Продуценты
(растения)
 неколич. потенциальная энергия неколич. потенциальная энергия
кинетическая энергия органических соединений
 солнечн. света колич. солнечн. света колич.
тепловая энергия
Консументы
(хищники, травоядные)
 потенциальная энергия потенциальная энергия
неколич. собственной протоплазмы
потенциальная энергия
соед. синтезинрованных
растениями  колич. колич.
тепловая энергия
Таким образом, все типы экосистем регулируются теми же основными законами, которые управляют и неживыми системами. Но есть и различия.
Еще в 1935 г. советский ученый Э.С.Бауэр в своей "Теоретической биологии" сформулировал три основные особенности живых систем.
1. Способность к самопроизвольному, без воздействия окружающей среды, изменению состояния.
2. Противодействие внешним силам, приводящее к изменению первоначального состояния окружающей среды.
3. Постоянная работа против уравновешивания с окружающей средой.
Первые две особенности встречаются и у других систем, а вот третья является отличительным признаком живых. Поэтому Бауэр назвал ее "всеобщим законом биологии", который имеет ясный термодинамический смысл - как в неживых системах устойчиво их равновесное состояние, так в живых устойчиво неравновесное.
Шредингер (1945 г.) тоже считает особенностью живых систем их неуравновешенность с окружающей средой, которая поддерживается непрерывным обменом открытой живой системы с окружающий средой едой, питьем, дыханием и т.д. Но обмен сам по себе ничего дать не может. Любой атом азота, кислорода, серы и т.п. также хорош, как и любой другой такого же рода. Может быть, целью обмена является поглощение энергии. Но ведь в зрелом организме содержание материи также постоянно, как и содержание энергии поэтому замена одного джоуля другим ничего не меняет. Более того, потребление пищи (энергии) взрослым организмом, как правило, значительно превышает потребности молодого, которому нужно интенсивно синтезировать собственную протоплазму. Значит, постоянный приток пищи (энергии) необходим живым системам не только для накопления энергии на черный день, либо для построения организма, т.е. для синтеза органических соединений, характерных для данного вида, и главным образом не для этого. Чтобы разобраться в этой проблеме рассмотрим, а как ведут себя неживые неравновесные системы. Если неживую неуравновешенную с окружающей средой систему изолировать, то всякое движение в ней скоро прекратится. В результате трения, теплопроводности, химических реакций и других самопроизвольных процессов потенциалы выравняются, система в целом угаснет и превратится в инертную массу материи, находящуюся в состоянии термодинамического равновесия, то есть максимальной энтропии. (Хороший пример - растворение кристаллика поваренной соли. На последнем примере удобно показать, что происходит с энтропией в самопроизвольных процессах. Кристаллик - упорядоченная ионная структура, где у каждого иона было определенное место; при растворении эта структура нарушилась, произошло разупорядочение структуры, энтропия увеличилась).
Таким образом, все, что происходит в природе, ведет к увеличению энтропии в той части мира, где это происходит, включая живые системы. Последние тоже непрерывно увеличивают свою энтропию, то есть производят положительную энтропию, и приближаются к опасному состоянию максимальной энтропии - смерти. Следовательно, неравновесное состояние живых систем (которые представляют собой чрезвычайно маловероятную структуру, обладающую очень низкой энтропией) поддерживается за счет извлечения ими из окружающей среды отрицательной энтропии - негоэнтропии. Назначение обмена - освободиться от производимой положительной энтропии и извлечь отрицательную. Но чем выше энтропия, тем больше беспорядок, и наоборот. Поэтому извлечение негоэнтропии есть "извлечение порядка", повышение упорядоченности системы, организма.
Есть два различных механизма, производящих упорядоченные явления: статический, создающий порядок из беспорядка; и механизм, создающий порядок из порядка низшего уровня. Закон сохранения энергии ничего не дает для их объяснения. Видимо, его надо искать на основе второго закона. Известно, что высшие животные питаются хорошо упорядоченными органическими соединениями. Использовав упорядоченность этих продуктов, животные возвращают в окружающую среду вещества в очень деградировавшей, неупорядоченной форме, там они усваиваются растениями. Для последних же мощным средством выработки отрицательной энтропии является солнечный свет, с помощью которого в хлорофилле происходит повышение упорядоченности деградировавших веществ - фотосинтез, и цикл повторяется. Это единственный на Земле естественный, самопроизвольный процесс, в котором энтропия уменьшается - за счет затраты даровой солнечной энергии. В соответствии со вторым законом, кинетическая энергия света превращается в потенциальную энергию связи органических соединений не количественно. Да, коэффициент этого перехода много меньше 100 %. Но энергия света достается даром! Нам все равно, с каким КПД ее будут расходовать растения, пусть он будет даже очень мал. Главное, растения и все "живое" обладают тайнами механизмов концентрирования и диссипирования энергии.
Таким образом, важнейшая термодинамическая характеристика организмов, экосистем и биосферы в целом - способность создавать и поддерживать высокую степень внутренней упорядоченности, то есть неуравновешенное состояние с низкой энтропией (с окружающей средой, но неустойчивое равновесие для самого индивида). Для поддержания внутренней упорядоченности в системе, находящейся при температуре выше абсолютного нуля, когда существует тепловое движение атомов и молекул, необходима постоянная работа по откачиванию "неупорядоченности". Эта работа предполагает постоянно действующий источник энергии и наличие хорошо развитых "диссипативных структур" у самой системы. Низкая энтропия достигается постоянным и эффективным рассеянием легко используемой концентрированной энергии (например, энергии света, горючего, пищи) и превращением ее в энергию, используемую с трудом (например, в тепловую). Дыхание высокоупорядоченной биомассы можно рассматривать как диссипативную структуру экосистемы. Это затрата энергии на поддержание жизнедеятельности.
Итак, экосистемы и организмы представляют собой открытые неравновесные термодинамические системы, постоянно обменивающиеся с окружающей средой энергией и веществом, уменьшая этим энтропию
внутри себя, но увеличивая энтропию вовне в согласии с законами термодинамики.
Энергетические характеристики среды.
Для описания "поведения" энергии в экосистемах подходит понятие "поток энергии", поскольку в отличие от циклического движения веществ превращения энергии идут в одном направлении. Энергия, однажды использованная каким-либо организмом, превращается в тепло и утрачивается для экосистемы. Энергия может накапливаться, сберегаться (т.е. преобразовываться в более эффективные формы) и передаваться из одной части системы в другую, но она не может быть снова пущена в дело, как вода и минеральные вещества. Живые замкнутые термодинамические системы невозможны. Каждый живой компонент, будь это организм или экосистема, должен получить от своей среды на входе постоянный приток энергии.
Основной источник практически всей энергии на Земле
- Солнце. На биосферу из космоса падает солнечный свет с энергией 2 кал·см-2
·мин-1
. Проходя через атмосферу, излучение экспоненциально ослабляется атмосферными газами и пылью и в ясный летний полдень до поверхности Земли может дойти не более 67 % его энергии. Степень этого ослабления зависит от длины волны (частоты) света. Ультрафиолетовое излучение с длиной волны менее 300 нм, почти не проходит через озоновый слой, имеющийся в атмосфере на высоте около 25 км, и это очень удачно, поскольку такое излучение летально для незащищенной протоплазмы. Излучение в оптической области (видимый свет) ослабляется равномерно, а инфракрасное излучение поглощается в атмосфере неодинаково в зависимости от длины волны. Лучистая энергия, достигающая земной поверхности в ясный день, состоит примерно на 10 % из ультрафиолетового излучения, на 45 % - из видимого света и на 45 % - из инфракрасного. Меньше всего при прохождении через плотные облака и воду ослабляется видимый свет. Следовательно, фотосинтез, нуждающийся именно в видимом свате, может идти и в пасмурные дни, и под слоем чистой воды некоторой толщины (10-20 м). Поступление солнечной энергии к автотрофному слою экосистемы за день обычно варьирует от 100 до 800 кал·см-2
, в среднем составляя около 300 - 400 кал·см-2
(3000 - 4000 ккал·м-2
) или 1,1 - 1,5 млн. ккал·м-2
·год-1
Растительность сильно поглощает синие и красные лучи, а также дальнее инфракрасное излучение; зеленый свет поглощается не так сильно (мы видим в отраженном свете, поэтому растительность для нас преимущественно зеленых оттенков); ближнее инфракрасное излучение - очень слабо. Тенистая прохлада создается в лесу благодаря тому, что листва поглощает много видимого и дальнего инфракрасного излучения. Синий и красный свет (400-500 и 600-700 нм соответственно) поглощаются особенно сильно хлорофиллом, а энергия дальнего инфракрасного излучения - водой, содержащейся в листьях, и окружающими их водяными парами. Таким образом, зеленые растения эффективно поглощают синий и красный свет, наиболее важный для фотосинтеза. Как бы отбрасывая ближнее ИК-излучение, несущее основную часть солнечной тепловой энергии, листья наземных растений избегают опасного перегрева. Кроме того, листья охлаждаются за счет испарения, а водные растения, разумеется, охлаждаются водой.
Таблица 1.
Рассеяние энергии солнечного излучения
( в % от годового поступления в биосферу). (По Hulbert, 1971.)
| % |
Отражрается
Прямо превращается в тепло
Испарение, осадки
Ветер, волны, течения
Фотосинтез
|
30
46
23
0,2
0,8
|
Энергия приливов - около 0,0017% солнечной энергии.
Тепло Земли - около 0,5% солнечной энергии.
|
Судьба солнечной энергии, поступающей в биосферу, кратко рассмотрена в табл. 1. Хотя всего лишь около 1 % энергии переходит в пищу и другую биомассу, те примерно 70 %, которые преобразуются в тепло, уходят на испарение, осадки, ветер и т.д., не теряются зря, т.к. эта энергия поддерживает температуру и приводит к действие системы погоды и круговорот воды, необходимые для жизни на Земле.
Другой энергетический компонент среды обитания - тепловое излучение. Оно исходит от всех поверхностей и тел, температура которых выше абсолютного нуля. Это не только почва, вода и растения, но и облака, излучающие вниз, на экосистемы, значительные количества тепловой энергии. Потоки длинноволнового излучения, разумеется, распространяются беспрестанно и во всех направлениях, а солнечный компонент (прямая солнечная радиация) имеет четкую направленность и поступает только днем. Следовательно, количество тепловой энергии, получаемой со всех сторон за сутки летом животным на открытом пространстве или листом растения, может в несколько раз превышать направленное прямо вниз излучение Солнца. Кроме того, тепловая энергия поглощается биомассой полнее, чем солнечное излучение.
Условия существования организмов (температура, скорость испарения воды, движения воздуха и воды) определяются общим потоком излучения, но для продуктивности экосистемы и для круговорота биогенных элементов в ней важнее суммарное прямое солнечное излучение, попадающее на автотрофный ярус экосистемы, т.е. солнечная энергия, получаемая зелеными растениями за недели, месяцы, весь год. Этот приток первичной энергии приводит в действие все биологические системы. Итак, мы подошли к концепции продуктивности.
Лекция 4.
Энергия и продуктивность.
1. Концепция продуктивности.
2. Энергетическая классификация систем.
3. Концепция градиента от субсидии до стресса.
Концепция продуктивности
Первичная продуктивность экологической системы, сообщества или любой их части определяется как скорость, с которой лучистая энергия усваивается организмами-продуцентами (главным образом зелеными растениями) в процессе фотосинтеза и хемосинтеза, накапливаясь в форме органических веществ.
В процессе производства органического вещества следует выделить четыре последовательных уровня, или ступени.
1. Валовая первичная продуктивность - это общая скорость фотосинтеза, включая те органические вещества, которые за время измерений были израсходованы на дыхание ("валовый фотосинтез", "общая ассимиляция").
2. Чистая первичная продуктивность - скорость накопления органического вещества в растительных тканях за вычетом того органического вещества, которое использовалось при дыхании растений за изучаемый период ("наблюдаемый фотосинтез", "чистая ассимиляция").
3. Чистая продуктивность сообщества - скорость накопления органического вещества, не потребленного гетеротрофами (т.е. чистая первичная продукция минус потребление гетеротрофами) за учетный период.
4. Вторичная продуктивность - скорость накопления энергии на уровнях консументов.
Высокие скорости продуцирования наблюдаются в естественных и искусственных экосистемах там, где физические факторы благоприятны, и особенно при поступлении дополнительной энергии извне, уменьшающем собственные затраты на поддержание жизнедеятельности. Такая дополнительная энергия может поступать в разной форме: в тропическом дождевом лесу - в форме работы ветра и дождя, в эстуарии - в форме энергии прилива, на возделываемом поле - в форме энергии ископаемого топлива и работы, совершаемой человеком или животным. Оценивая продуктивность экосистемы, необходимо учитывать как утечки энергии, связанные со сбором урожая, загрязнением среды, неблагоприятными климатическими условиями и другими типами стрессовых воздействий, способствующих отведению энергии от процесса продукции, так и поступления энергии, которые увеличивают продуктивность, компенсируя потери тепла при дыхании (при "откачивании неупорядоченности") необходимые для поддержания биологической структуры.
Всякий источник энергии, уменьшающий затраты на самоподдержание экосистемы и увеличивающий ту долю энергии, которая может перейти в продукцию, называется вспомогательным потоком энергии или энергетической субсидией.
Энергетическая классификация систем
.
Источник и качество доступной энергии в той или иной степени определяют видовой состав и численность организмов, характер функциональных процессов, протекающих в экосистеме, и процессов ее развития, а также образ жизни человека. Энергия - общий знаменатель и исходная движущая сила всех экосистем, как сконструированных человеком, так и природных, следовательно, логично принять энергию за основу для "первичной классификации экосистем. Удобно выделить на этой основе четыре фундаментальных типа экосистем.
1. природные, движимые Солнцем, несубсидируемые;
2. природные, движимые Солнцем, субсидируемые другими естественными источниками энергии;
3. движимые Солнцем и субсидируемые человеком;
4. индустриально-городские, движимые топливом (ископаемым, другим органическим или ядерным).
Энергетическая классификация основана на свойствах среды на входе, она коренным образом отличается от биомной классификации, основанной на внутренней структуре экосистем, но вместе с тем и дополняет ее.
Природные системы, в основном или полностью зависящие от прямого солнечного излучения, можно назвать движимыми Солнцем несубсидируемыми экосистемами. Они совсем или почти не получают дополнительной энергии, помимо солнечного света. К числу таких экосистем можно отнести открытие океаны, крупные участки горных лесов, грасленды и большие глубокие озера. Часто на них накладываются и другие ограничения, например, нехватка элементов питания и воды. Поэтому хотя экосистемы этой обширной группы весьма различны, все они получают мало энергии (от 1000 - 10000 ккал·м-2
·год-1
) и имеют низкую продуктивность или способность выполнять работу. Организмы, живущие в таких системах, выработали замечательные адаптации к существованию на скудном пайке энергии и других ресурсов и к эффективному их использованию.
Хотя мощность природных экосистем, относящихся к первой категории, не очень впечатляет, и они не способны поддерживать высокую плотность населения, тем не менее такие экосистемы крайне важны, так как занимают огромные площади (одни лишь океаны - до 70 % площади земного шара). Весь комплекс движимых Солнцем природных экосистем крайне важен для человека, это по сути дела основной "модуль жизнеобеспечения", гомеостат, стабилизирующий и поддерживающий условия на "космическом корабле", имя которому Земля; именно здесь ежедневно очищаются большие объемы воздуха, возвращается в оборот вода, формируются климатические условия, измеряются крайности погоды и выполняется множество других полезных функций.
Если помимо солнечного света могут быть использованы какие-то дополнительные источники энергии, плотность мощности может быть значительно повышена, порой даже на порядок величины (10000 - 40000 ккал·м-2
·год-1
). В этом случае несолнечная энергия частично заменяет солнечную, сокращая расходы на самоподдержание системы и высвобождает солнечную энергию на производство органических веществ. Источники дополнительной энергии могут быть как естественными, так и искусственными. Для простоты классификации выделены категории: движимые Солнцем экосистемы с естественными и с искусственными энергетическими субсидиями.
Вспомогательная энергия, увеличивающая продуктивность, может поступать в самых разнообразных формах, например, в тропическом дождевом лесу - в форме ветра и дождя, в небольшом озере - в форме потока воды из ручья, или поступающих с площади водосбора органических веществ и минеральных элементов. Прибрежная часть эстуария - хороший пример природной экосистемы с дополнительной энергией приливов, прибоя и течений. Поскольку приливы и течения воды способствуют более быстрому круговороту минеральных элементов питания и перемещению пищи и отходов, организмы в эстуарии могут, так сказать, сконцентрировать свои усилия на более эффективном превращении энергии Солнца в органическое вещество.
Человек также давно научился изменять природу и использовать вспомогательные источники энергии для получения прямой выгоды, а его умение не только увеличивать продуктивность, но и направлять эту продуктивность на производство пищевых и волокнистых материалов, легко собираемых, перерабатываемых и используемых, постоянно растет. Наземные и водные агроэкосистемы - основные примеры систем движимых Солнцем и субсидируемых человеком. Высокая продуктивность поддерживается большими поступлениями энергии топлива (а при более примитивных системах сельского хозяйства - мышечных усилий человека и животных). Эта энергия тратится на возделывание, орошение и удобрение, селекцию и борьбу с вредителями. Самое продуктивное сельское хозяйство находится примерно на уровне самых продуктивных природных экосистем: по-видимому, верхний предел для любой постоянной, длительно функционирующей системы, основанной на фотосинтезе, составляет примерно 50000 ккал·м-2
·год-1
. Действительное различие между природными и искусственными экосистемами состоит лишь в распределении этого потока энергии. Человек старается направить как можно больше энергии на производство продуктов питания, которые он может немедленно использовать, а природа обычно распределяет продукты фотосинтеза между многими видами и веществами и накапливает энергию "на черный день", это, так называемая, стратегия повышения разнообразия в целях выживания.
В экосистеме, движимой топливом высококонцентрированная потенциальная энергия топлива не просто дополняет, а заменяет солнечную энергию. При современных методах ведения городского хозяйства солнечная энергия в самом городе не только не используется, но становится дорогостоящей помехой, так как она нагревает бетон и способствует образованию смога. Важное свойство экосистем, движимых горючим - огромная потребность в энергии плотно населенных индустриально-городских районов, она по меньшей мере на 2-3 порядка больше того потока энергии, который поддерживает жизнь в естественных условиях. Килокалории энергии, ежегодно протекающие через квадратный метр индустриального города, исчисляется уже не тысячами, а миллионами ( 100000 - 3000000 ккал·м-2
·год-1
). Вот почему множество людей могут жить на небольшой территории.
Рассматривая общую концепцию энергетических субсидий надо сделать еще одно замечание, фактор, который при одних условиях среды или при одном уровне поступлений увеличивает продуктивность, при других условиях среды или другом уровне поступлений может способствовать утечкам энергии, уменьшая продуктивность. Слишком много хорошего также вредит системе, как и слишком мало. Например, некоторые виды загрязнений - обработанные сточные воды, могут в зависимости от объема и периодичности сброса оказаться либо благоприятным фактором, либо источником стресса. Если обработанные сточные воды попадают в экосистему с постоянной умеренной скоростью, то они, доставляя в систему микроэлементы, могут способствовать повышению продуктивности, однако массовый их сброс через нерегулярные промежутки времени может почти полностью уничтожить систему как биологическую единицу.
Концепция градиента от субсидии до стресса.
Концепция градиента от субсидии до стресса иллюстрируется рис. 2. Если нарушение на входе вредно, то реакция экосистемы должна быть отрицательной. Но если на вход поступают полезные вещества или энергия, то продуктивность (или другая мера эффективности) может увеличиться: такая экосистема и будет называться субсидируемой экосистемой. При увеличении поступления субсидий способность системы их усваивать может достичь предела; эффективность после этого снизится. Хорошая иллюстрация соотношений между величиной субсидий и стрессовым воздействием - реакция системы на повышенное внесение азотных удобрений.
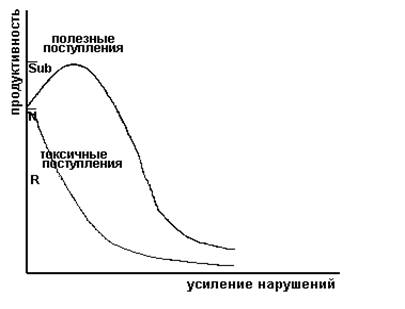
Рис.2. Иллюстрация концепции градиента от субсидии до стресса.
Лекция 5.
Пищевые цепи, пищевые сети и трофические уровни.
1. Принципы организации и типы пищевых цепей.
2. Компоненты "универсальной" модели экологического потока энергии.
3. Экологическая эффективность.
4. Концепция трофического уровня.
5. Концепция поддерживающей емкости.
Мы рассмотрели общий поток энергии, характеризующий среду экосистемы, рассмотрели качество энергии и основанную на энергии классификацию экосистем. Настало время обратиться к изучению той части общего потока энергии, которая проходит через живые компоненты экосистемы.
Перенос энергии пищи от ее источника - автотрофов (растений) через ряд организмов, происходящий путем поедания одних организмов другими, называется пищевой цепью. Пищевые цепи можно разделить на два типа: пастбищная цепь, которая начинается с зеленого растения и идет далее к пасущимся растительноядным животным (т.е. к организмам, поедающим живые растительные клетки или ткани) и к хищникам (организмам, поедающим животных); и детритная цепь, которая от мертвого органического вещества идет к микроорганизмам, а затем к детритофагам и к их хищникам. Пищевые цепи не изолированы одна от другой, а тесно переплетаются друг с другом, образуя, так называемые, пищевые сети.
В сложных природных сообществах организмы, получающие свою энергию от Солнца через одинаковое число ступеней, считаются принадлежащими к одному трофическому уровню. Первый трофический уровень представлен первичными продуцентами или автотрофами; к ним относятся зеленые растения, которые способны использовать солнечный свет для образования химических соединений, богатых энергией. Первичные продуценты - это весьма важная часть сообщества, потому что практически все остальные организмы, входящие в его состав, прямо или косвенно зависят от снабжения энергией, запасенной растениями. Помимо первичных продуцентов в состав сообщества входят гетеротрофы, которые представлены консументами и деструкторами. Второй трофический уровень образуют растительноядные животные, называемые первичными консументами. Плотоядных, которые питаются растительно-травоядными называют вторичными консументами или первичными хищниками; они занимают третий трофический уровень. Хищники, питающиеся первичными хищниками, в свою очередь, образуют четвертый трофический уровень и называются третичными консументами или вторичными хищниками. Точно также животные, потребляющие вторичных хищников называются четвертичными консументами или третичными хищниками, они находятся на пятом трофическом уровне и т.д.
Эта трофическая классификация относится к функциям, а не к видам как таковым. Поскольку многие животные всеядны и питаются как растениями, так и животными, т.е. одновременно получают энергию с нескольких разных трофических уровней, их невозможно отнести к определенному уровню. Принято считать, что такие организмы представляют сразу несколько трофических уровней, а их участие в каждом уровне пропорционально составу их диеты.
В исследованиях структуры сообществ понятие трофического уровня оказалось чрезвычайно полезной абстракцией. Оно облегчает изучение потока вещества и энергии через сообщество и подчеркивает различия между взаимодействиями, которые протекают внутри трофических уровней и между ними.
Принцип организации пищевых цепей и действия двух законов термодинамики можно уяснить, рассмотрев схему переноса энергии на рис. 3. На этой схеме четырехугольники изображают трофические уровни, "трубы" - потоки энергии от каждого уровня или к нему. Как требует первый закон термодинамики, приток энергии уравновешивается ее оттоком, и каждый перенос энергии сопровождается ее рассеянием в форме недоступной для использования тепловой энергии (при дыхании), как того требует второй закон.
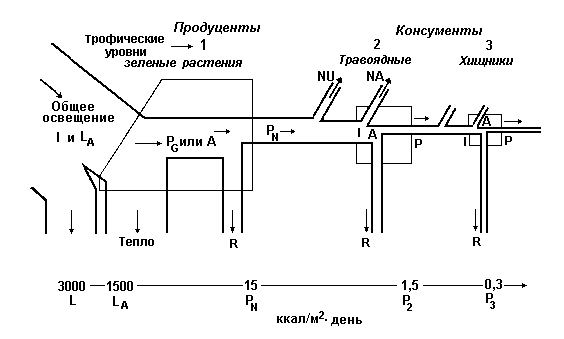
Рис.3. Упрощенная схема потока энергии, показывающая три трофическиих уровня (I, II и III ) в линейной пищевой цепи ( E.Odum, 1963.)
I - общее поступление энергии; LA
- свет, поглощаемый растительным покровом; PG
- валовая первичная продуктивность ; А - общая ассимиляция; PN
- чистая первичная продукция; P2-3
- вторичная продукция (консументов); NU- неиспользуемая (накапливаемая или экспортируемая энергия); NA- неассимилированная консументами (выделенная с экскрементами) энергия; R - дыхание.
Представленная схема потоков энергии на трех трофических уровнях сильно упрощена. Но она позволяет ввести принятые в литературе обозначения разных потоков и ясно показывает, что на каждом последующем уровне поток энергии сильно уменьшается независимо от того, рассматривается ли общий поток (I - общий поток энергии и А - общая ассимиляция) или компоненты Р (продуктивность биомассы)и Р (дыхание). Показано, что на первом трофическом уровне поглощается около 50 % падающего света, а превращается в энергию пищи всего 1 % поглощенной энергии, а также "двойной метаболизм" продуцентов (т.е. валовая и чистая продукция). Вторичная продуктивность ( Р ) на каждом последующем трофическом уровне консументов составляет около 10 % предыдущей , хотя на уровне хищников эффективность может быть выше, скажем 20 %. Если питательная ценность источника энергии велика (например, продукт фотосинтеза, извлекаемый или выделяемой прямо из растительных тканей), то эффективность переноса энергии может быть гораздо выше. Но поскольку и растения, и животные производят, много трудноперевариваемого вещества (целлюлоза, лигнин, хитин), а также химические ингибиторы, препятствующие поеданию различными консументами, средняя эффективность переноса энергии между трофическими уровнями в целом составляет 20 % и менее.
Обратимся теперь к основным компонентам модели потока энергии. На рисунке 4 представлена модель, которую можно назвать "универсальной", поскольку она приложима к любому живому компоненту системы, будь то растение, животное, микроорганизм, особь, популяция или трофическая группа. Соединенные между собой, такие графические модели могут отразить, как уже показано (рис. 3) пищевые цепи или биоэнергетику экосистемы в целом.
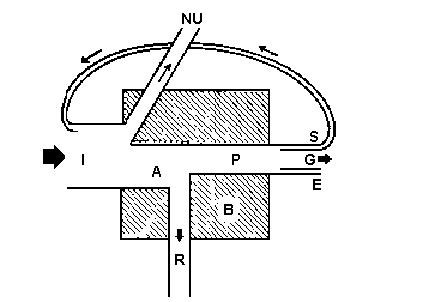
Рис. 4. Компоненты "универсальной" модели экологического потока энергии. (E. Odum, 1968.)
На рис. 4 квадрат, обозначенный буквой В, изображает живую структуру или биомассу основного компонента модели. Желательно выражать биомассу в калориях, чтобы выявить связь между размерами потока энергии и одномоментной или средней биомассой. Общее поступление энергии - источник, обозначено буквой I. Для облигатных автотрофов это свет, для облигатных гетеротрофов - органическая пища. Некоторые водоросли и бактерии могут использовать оба источника энергии, а многие из них нуждаются в определенном соотношении этих источников. Сходная ситуация наблюдается у лишайников и тех беспозвоночных животных, в которых содержатся водоросли-мутуалисты. В таких случаях канал притока на схеме потоков энергии можно разделить пропорционально использованию различных источников энергии; либо можно разделить биомассу на отдельные резервуары. Тогда каждый резервуар (квадрат) будет соответствовать одному энергетическому (трофическому) уровню.
Не вся энергия, поступающая в биомассу, подвергается превращению. Часть ее может просто пройти через биологическую структуру, например пища может пройти через пищеварительный тракт, не включаясь в метаболизм, а часть энергии света проходит через растительность не усваиваясь. Эта часть энергии обозначается через NA. Используемая, или ассимилируемая, часть на схеме обозначена буквой А. Отношение А и I, т.е. эффективность ассимиляции, широко варьирует. Оно может быть очень мало, как в случае усвоения света растениями (< 1 %) или ассимиляции пищи у животных - детритофагов, или же очень велико, как в случае животных или бактерий, питающихся высококалорийной пищей, например сахарами и аминокислотами. Что указывает на важность такого параметра, как качество ресурса.
У автотрофов ассимилированная энергия (А) - это валовая продукция или валовый фотосинтез. Аналогичный компонент (А) у гетеротрофов - это пища, продуцированная кем-то другим. У высших животных величину А часто называют "метаболизированной энергией".
Ключевая особенность этой модели - разделение ассимилированной энергии на компоненты Р и R. Та часть фиксированной энергии А, которая окисляется и теряется в форме тепла, называется дыханием (R), а та часть, которая превращается в новое или принадлежащее другому виду органическое вещество, называется продукцией (Р). У растений это чистая продукция, у животных - вторичная продукция. Компонент Р - это энергия, доступная следующему трофическому уровню, в противоположность компоненту NU , который доступен и на данном трофическом уровне.
Отношения R/Р и R/B широко варьируют. Они имеют важное экологическое значение. В экосистеме отношение общего дыхания сообщества к его суммарной биомассе (R/B ) можно рассматривать как отношение затрат энергии на поддержание жизнедеятельности к энергии, заключенной в структуре, или как меру термодинамической упорядоченности. Это "соотношение Шредингера" служит мерой экологического оборота. Если выразить R и B в калориях и отнести их к абсолютной температуре, то отношение R/B становится отношением прироста энтропии, связанного с поддержанием структуры, к энтропии упорядоченной части. В целом часть энергии, идущая на дыхание, или на поддержание структуры, велика в популяциях крупных организмов (люди, деревья) и в сообществах с большой биомассой на корню. При стрессовых воздействиях на систему R возрастает.
Величина P, напротив, сравнительно велика в активных популяциях мелких организмов, например, бактерий или водорослей, в молодых, быстро растущих сообществах и в системах, получающих энергетические дотации. Продукция может принимать различные формы. Три ее типа указаны на рисунке 4. (G - рост, или увеличение, биомассы, Е - ассимилированное органическое вещество, выделяемое с экскрементами, секретируемое или извлекаемое, S- запас (например, жировые накопления, которые могут быть использованы позже). Обратный поток S можно назвать и "рабочей петлей", поскольку это та часть продукции, которая используется для обеспечения энергией в будущем (например, хищник использует энергию запасенных веществ, чтобы найти новую жертву, энергия экскрементов или выделений травоядного животного удобряет почву, поддерживая жизнь растений, которыми это животное питается).
Значительный интерес для эколога представляют отношения величин энергетического потока в разных точках пищевой цепи. Выраженные в процентах, эти отношения часто называют экологической эффективностью. В табл. 2 приведены некоторые из этих отношений и даны определения, в основе которых лежат представления об энергетических потоках. По большей части эти отношения могут использоваться применительно и к составляющим популяциям, и к целым трофическим уровням.
Таблица 2.
Различные типы экологической эффективности
| Отношение |
Название и объяснение |
It
/ It-1
At
/ At-1
Pt
/ Pt-1
It
/ Pt-1
или At
/ Pt-1
Pt
/ At
Pt
/ It
At
/ It
|
А. Отношения между трофическими уровнями
Эффективность поглощения энергии трофическим уровнем
Эффективность ассимиляции трофического уровня
Эффективность продукции трофического уровня
Эффективность использования
Б. Отношения внутри трофических уровней
Эффективность роста тканей или продукции
Экологическая эффективность роста
Эффективность ассимиляции
|
Уже говорилось об эффективности переноса энергии между трофическими уровнями; PG
/L сбычно равно 1-5 %, PG
/LA
равно 2-10 %, и эффективность переноса продукции между вторичными трофическими уровнями составляет обычно 10-2О %.
Поскольку у теплокровных животных (эндотермов), постоянно поддерживающих высокую температуру тела, доля ассимилированной энергии, которая должна идти на дыхание, как минимум в 10 раз больше, чем у холоднокровных (экзотермов), у первых должна быть ниже эффективность продукции Р/A. Поэтому в "беспозвоночной" пищевой цепи эффективность переноса энергии между трофическими уровнями должна быть выше, чем в цепи с участием млекопитающих.
Как правило, эффективность роста тканей и эффективность ассимиляции у животных связаны обратной зависимостью. Для травоядных животных обычно характерна более высокая эффективность Р/А , но более ниакая А/I, чем для хищников.
Многих озадачивала очень низкая первичная эффективность, характерная для интактных природных систем, в сравнении с высокими КПД электромоторов и других двигателей. Это привело к мысли о необходимости серьезно рассмотреть возможности увеличения эффективности процессов, происходящих в природе. На самом деде, долгоживущие, крупномасштабные экосистемы нельзя приравнивать в этом отношении к недолговечным механическим системам. Во-первых, в живых системах много "горючего" затрачивается на "ремонт" и самоподдержание, а при расчете КПД двигателей не учитываются амортизация и расходы энергии на ремонт. Во-вторых, в определенных условиях быстрый рост может иметь большее значение для выживания, чем максимальная эффективность использования "горючего". (Простая аналогия: автомобилисту может быть важнее быстро достичь пункта назначения, развив высокую скорость, чем с максимальной эффективностью использовать бензин, придерживаясь малой скорости).
Инженерам важно понять, что любое повышение эффективности биологической системы обернется увеличением затрат на ее поддержание. По мере того, как размеры и сложность системы увеличиваются, пропорционально, но еще быстрее увеличивается энергетическая стоимость поддержания структуры и функции системы. При удвоении размеров системы, как правило, более чем вдвое увеличивается количество энергии, которая должна отводиться на уменьшение энтропии, связанной с необходимостью сохранения структурной и функциональной сложности. При увеличении размера и сложности системы проявляется закон увеличения отдачи, или экономия, связанная с масштабами, например, возрастает качество и устойчивость системы к нарушающим воздействиям. Однако при этом наблюдается также уменьшение отдачи или, расходы, связанные с масштабами, что объясняется увеличением стоимости откачивания неупорядоченности. Такое уменьшение отдачи внутренне присуще большим и сложным системам. Его можно сгладить, улучшив конструкцию системы, что увеличивает эффективность превращений энергии, но полностью устранить невозможно.
Являются ли такого рода расходы, связанные с масштабами, неотъемлемым свойством экосистем, неизвестно, но по крайней мере часть возрастающей платы за сложность компенсируется преимуществами (экономия, связанная с масштабами). Метаболизм на единицу массы уменьшается с увеличением массы организма или биомассы леса, так что на единицу потока энергии удается поддерживать больше структурных образований. Добавочные функциональные цепи и петли обратной связи могут увеличить эффективность использования энергии и повторного использования веществ и могут повысить устойчивость или упругость по отношению к нарушающим воздействиям. Возможность проявления эмерджентных свойств, в том числе симбиоза между организмами, может увеличить общую эффективность.
Но какие бы приспособления не возникали в системе, общая энтропия быстро увеличивается с ростом размеров. В результате все большая и большая доля общего потока энергии должна отклоняться на дыхание, связанное с поддержанием системы, в связи с чем все меньшая доля остается для нового роста. Когда расходы энергии на поддержание уравниваются с количеством доступной энергии, дальнейший рост прекращается, достигается теоретическая максимальная поддерживающая емкость.
Концепцию поддерживающей емкости можно объяснить с помощью диаграмм типа показанных на рис. 5.
Поддерживающая
емкость среды
|
|
                   Рис.5. Поддерживающая способность среды и ее связь с сигмоидным ростом популяции.
К - максимальная плотность, которая может сохраняться на данном пространстве и данной базе ресурсов;
I - уровень, при котором скорость роста максимальна. Это теоретический оптимум, при котором устойчивый урожай популяции максимален;
Диапазон от I до К - соответствует безопасной или желательной плотности.
Рост размеров и сложности популяций, а также целых экосистем часто идет по S -образной или сигмоидной кривой. Обратим внимание на две точки на кривой роста: К - верхняя асимптота, соответствует максимальной поддерживающей емкости, т.е. количеству биомассы, которая может поддерживаться при данных условиях, а I - точка перегиба, где скорость роста максимальна. Уровень I называют максимальным постоянным выходом, либо оптимальной поддерживающей емкостью, способной сохраняться долгое время несмотря на капризы среды. Согласно теории сложности, собранная биомасса будет быстрее всего восстанавливаться на этом уровне.
Сложность поддержания максимального уровня (К-уровня) в колеблющихся условиях реального мира состоит в том, что неизбежны отклонения от этого уровня: превышения вследствие инерции роста популяций и снижения (хотя бы временные) вследствие периодического уменьшения доступных ресурсов (например, при засухе). Когда уровень становится выше максимального и энтропия превосходит способность системы рассеивать ее, наблюдается уменьшение размеров, или "крушение". Если при этом пострадала производительная способность среды, сам уровень К может временно снизиться (до уровня К¢). Безопасный допуск уровня максимальной поддерживающей емкости очень невелик. С точки зрения долгосрочной безопасности и стабильности уровень желательной поддерживающей емкости находится где-то между К и I. Это диапазон поддерживающей емкости .
Мировая проблема снабжения человечества продуктами питания заключается в том, что потребность в них сейчас приближается к максимальной производительной способности Земли при существующих технологических, политических, экономических и распределительных ограничениях. Любое сильное стрессовое воздействие, например, война, засуха или болезнь, которое сократит урожаи хотя бы на год, означает серьезное недоедание или голод для миллионов, еле сводящих концы с концами.
Лекция 6.
Трофическая структура и трофическая функция экосистемы.
1. Метаболизм и размеры особей.
2. Трофическая структура и экологические пирамиды.
3. Концентрация токсичных соединений при прохождениипо пищевым цепям.
Метаболизм и размеры особей.
Размер урожая биомассы на корню (выраженный в общей калорийности или в общей сухой массе всех организмов, присутствующих в данный момент времени), который может поддерживаться постоянным потоком энергии через пищевую цепь, в значительной степени зависит от размера особей. Чем меньше организм, тем выше его удельный метаболизм (т.е. метаболизм, обмен веществ в пересчете на 1 г или 1 кал, биомассы) и тем меньше биомасса, которая может поддерживаться на данном трофическом уровне. Так "урожай" бактерий, имеющихся в любой данный момент, будет гораздо ниже "урожая" рыбы или млекопитающих, хотя эти группы, возможно, используют одинаковое количество энергии.
Различные теории, объясняющие эту тенденцию, особое внимание обращают на роль процессов диффузии; действительно у крупных организмов площадь поверхности, через которую может идти диффузия, на грамм массы меньше, чем у мелких. Однако общепринятого объяснения связи между размером и метаболизмом пока нет.
Следует помнить, что с увеличением размеров снижается не общий метаболизм особи, а удельный. Взрослому человеку требуется пищи больше, чем маленькому ребенку, но на 1 кг массы взрослый потребляет меньше пищи.
Трофическая структура и экологические пирамиды
.
В результате взаимодействия энергетических явлений в пищевых цепях (потерь энергии при каждом переносе) и такого фактора, как зависимость метаболизма от размеров особи, каждое сообщество приобретает определенную трофическую структуру, которая часто служит характеристикой экосистемы (озера, леса, пастбища и т.д.). Трофическую структуру можно измерить и выразить либо урожаем на корню на единицу площади, либо количеством энергии, фиксируемой на единицу площади за единицу времени на последовательных трофических уровнях. Трофическую структуру и трофическую функцию можно изобразить графически в виде экологических пирамид. Основанием этих пирамид служит первый трофический уровень - уровень продуцентов, а последующие уровни образуют этажи и вершину пирамиды. Экологические пирамиды можно отнести к трем основным типам:
1) пирамида численностей, отражающая численность отдельных организмов;
2) пирамида биомассы, характеризующая общую сухую массу, калорийность или другую меру общего количества живого вещества ;
3) пирамида энергии, показывающая величину потока энергии и (или) "продуктивность" на последовательных трофических уровнях.
Пирамиды численности и массы могут быть обращенными, или частично обращенными, т.е. основание может быть меньше, чем один или несколько верхних этажей. Так бывает, когда средние размеры продуцентов меньше размеров консументов. Можно сформулировать некое "экологическое правило": данные по численности приводят к переоценке значения мелких организмов, а данные по биомассе - к переоценке роли крупных организмов. Следовательно, эти критерии непригодны для сравнения функциональной роли популяций, сильно различающихся по отношению интенсивности метаболизма к размеру особей, хотя, как правило, биомасса все же более надежный критерий, нежели численность.
Из трех типов экологических пирамид пирамида энергии дает наиболее полное представление о функциональной организации сообществ. Число и масса организмов, которых может поддерживать какой-либо уровень в тех или иных условиях, зависит не от количества фиксированной энергии, имеющейся в данное время на предыдущем уровне, а от скорости продуцирования пищи. В противоположность пирамидам численностей и биомассы, отражающим статику системы (т.е. характеризующим количество организмов в данный момент), пирамида энергии отражает картину скоростей прохождения массы пищи через пищевую цепь. На форму этой пирамиды не влияют изменения размеров и интенсивности метаболизма особей, и если учтены все источники энергии, то пирамида всегда будет иметь "правильную форму", как это диктуется вторым законом термодинамики. Концепция потока энергии не только позволяет сравнивать экосистемы между собой, но и дает средство для оценки относительной роли популяций в их биотических сообществах.
Многие исследователи считают, что трофическая структура - фундаментальное свойство каждого сообщества, ибо после острого нарушения его трофическая структура возвращается к равновесию независимо от видового состава и быстрее его, т.е. трофическая структура обладает способностью восстанавливаться. И только если экосистема постоянно находится под стрессовым воздействием, трофическая структура может измениться по мере приспособления биотических компонентов экосистемы к хроническим нарушениям.
Концентрация токсичных соединений при прохождении по пищевым цепям
Распределение энергии не единственный количественный параметр, на который влияют события, происходящие в пищевых цепях. Некоторые вещества по мере продвижения по цепи не рассеиваются, а наоборот, накапливаются. Это , так называемое, концентрирование в пищевой цепи, нагляднее всего демонстрируют некоторые устойчивые радионуклиды и пестициды. Так было обнаружено, что коэффициент накопления (соотношение количества вещества в тканях и в окружающей среде) радиоактивного фосфора равен 2 миллионам. Таким образом "безопасные выбросы" в окружающую среду могут стать крайне опасными для высших звеньев пищевой цепи, к которым относится и человек.
Подобный пример демонстрирует и ДДТ. Чтобы сократить численность комаров на Лонг-Айленде, болота много лет опыляли ДДТ. Специалисты по борьбе с насекомыми благоразумно не применяли таких концентраций, которые были бы непосредственно летальны для рыбы и других животных, но они не учли экологических процессов и длительного сохранения токсичных остатков ДДТ. Ядовитые остатки, адсорбированные на детрите, концентрировались в тканях детритофагов и мелких рыб и далее - в хищниках высшего порядка - таких, как рыбоядные птицы. Коэффициент концентрации для рыбоядных птиц составляет около 500 000.
Принцип биологического накопления надо учитывать при любых решениях, связанных с поступлением загрязнений в среду. Многие небиологические факторы, однако, могут уменьшать или увеличивать коэффициент концентрации. Так человек получает меньше ДДТ, чем рыбоядные птицы и частично это объясняется тем, что при обработке и варке пищи часть этого вещества удаляется. Хищная рыба же находится в особенно опасном положении, потому, что может получать ДДТ не только через пищу, но и прямо из воды через жабры.
Лекция 7.
Биосфера как глобальная экосистема.
1. Понятие о биосфере.
2. Живое вещество биосферы.
3. Геохимическая работа живого вещества.
Понятие о биосфере .
Представление о биосфере как общепланетарной оболочке, охватывающей толщу тропосферы, гидросферы, осадочных (и возможно гранитных) пород литосферы, в ходе всей геологической истории Земли; как глобальной единой системе Земли, где весь основной ход геохимических и энергетических превращений определяется жизнью, было разработано в трудах В.И. Вернадского. Вернадский впервые указал на активную преобразующую деятельность древних и современных организмов в изменении облика нашей планеты. Грандиозные масштабы этого процесса позволили ему развить учение о космической роли жизни в геологической истории Земли, что несомненно дает право считать его основателем учения о биосфере.
Биосферой Вернадский назвал ту область нашей планеты, в которой существует или когда-либо существовала жизнь и которая постоянно подвергается или подвергалась воздействию живых организмов.
Участие каждого отдельного организма в геологической истории Земли ничтожно мало. Однако живых существ на Земле бесконечно много, они обладают высоким потенциалом размножения, активно взаимодействуют со средой обитания и в конечном счете представляют в своей совокупности особый, глобальных масштабов фактор, преобразующий верхние оболочки Земли.
Значение организмов обусловлено их большим разнообразием, повсеместным распространением, длительностью существования в истории Земли, избирательным характером биохимической деятельности и исключительно высокой химической активностью по сравнению с другими компонентами природы.
Биосфера, таким образом, это та область Земли, которая охвачена влиянием живого вещества. С современных позиций биосферу рассматривают как наиболее крупную экосистему планеты, поддерживающую глобальный круговорот веществ.
Современная жизнь распространена в верхней части земной коры (литосфере), в нижних слоях воздушной оболочки Земли (атмосфере) и в водной оболочке Земли (гидросфере). Для обозначения совокупности всего живого на Земле вместе с его непосредственным окружением и ресурсами введем термин "современная биосфера" или "экосфера".
Экосфера непрерывной оболочкой одевает земной шар, а ее протяженность по вертикали меняется от долей метра - в областях чрезвычайно скудной жизни (арктические и антарктические пустыни) - до тысяч метров. Нижняя граница экосферы ограничена прежде всего температурой горных пород и подземных вод, которая постепенно возрастает с глубиной и на уровне 1,5 - 15 км уже превышает 100°С. Поэтому вглубь Земли живые организмы проникают на небольшое расстояние. Самая большая глубина, на которой в породах земной коры были обнаружены бактерии, составляет 4 км. В нефтяных месторождениях на глубине 2 - 2,5 км бактерии регистрируются в значительном количестве. В океане жизнь распространена до более значительных глубин и встречается даже на дне океанических впадин в 10 - 11 км от поверхности, так как температура там около 0°С. Однако по Вернадскому нижнюю границу биосферы следует проводить еще глубже. Постепенно накапливающиеся в океане гигантские толщи осадочных пород, происхождение которых связано с деятельностью живых существ - это тоже часть биосферы. В соответствии с динамическими процессами в земной коре осадочные породы постепенно вовлекаются в глубь ее, метаморфизируясь под действием высоких температуры и давления. Метаморфические породы земной коры, происходящие из осадочных, в конечном итоге также производные жизни.
Верхняя граница жизни в атмосфере определяется нарастанием с высотой ультрафиолетовой радиации. На высоте 25 - 27 км большую часть ультрафиолетового излучения Солнца поглощает находящийся здесь тонкий слой озона - озоновый экран. Все живое, поднимающееся выше защитного слоя озона, погибает. Атмосфера же над поверхностью Земли насыщена многообразными живыми организмами. Споры бактерий и грибов обнаруживают до высоты 20 - 22 км, но основная часть аэропланктона сосредоточена в слое до 1-1,5 км.
Хотя процессы жизнедеятельности современных организмов сосредоточены только в экосфере, влияние живого вещества (современного или существовавшего в прошлом) ощущается далеко за ее пределами. Именно поэтому биосфера Вернадского (как область существования всех былых экосфер) простирается далеко за пределы современной экосферы, охватывая по вертикали слой толщиной 40 - 50 км.
Приблизительная масса биосферы составляет 0,05% массы Земли, а ее объем 0,4% объема планеты.
Структура биосферы представляет собой сложную многокомпонентную систему - совокупность газообразной, жидкой, твердой и биологической организаций. Она характеризуется строгой организованностью, биологическим равновесием численности и взаимной адаптированностью составляющих ее организмов.
Вернадский подчеркивал, что биосферу нужно рассматривать как целостную геологическую оболочку Земли, весьма сложную саморегулирующуюся систему, состоящую из живого вещества и неживой материи.
Всю совокупность организмов на планете Вернадский назвал живым веществом.
Косное вещество, по Вернадскому, это совокупность тех веществ в биосфере, в образовании которых живые организмы не участвуют - т.е. горные породы магматического, неорганического происхождения, видоизмененные живыми организмами вещества космического происхождения, космическая пыль, метеориты.
Биогенное вещество создается и перерабатывается жизнью, совокупностями живых организмов. Это источник чрезвычайно мощной потенциальной энергии (каменный уголь, гумус почв, нефть, битумы, торф и т.п.). После образования биогенного вещества живые организмы в нем малодеятельны.
Особой категорией является биокосное вещество. Вернадский определял, что оно "создается в биосфере одновременно живыми организмами и косными процессами, представляя системы динамического равновесия тех и других". Организмы в биокосном веществе играют ведущую роль. Биокосное вещество планеты - это почвы, кора выветривания, все природные воды, свойства которых зависят от деятельности на Земле живого вещества.
Живое вещество биосферы
В качестве основных характеристик живого вещества биосферы В.И.Вернадский рассматривал суммарную массу, химический состав и энергию. Отметим наиболее важные аспекты сравнительного анализа этих характеристик живого вещества с подобными для остальных оболочек биосферы.
Суммарный химический состав живых организмов во многом отличается от состава атмосферы и литосферы. Он ближе к химическому составу гидросферы по абсолютному преобладанию атомов водорода и кислорода, но, в отличие от гидросферы, в организмах относительно велика доля углерода, кальция, азота, фосфора и серы.
Живое вещество в основном состоит из элементов, являющихся водными и воздушными мигрантами, т.е. образующих газообразные и растворимые соединения. Заслуживает внимания то обстоятельство, что 99,9% массы живых организмов приходится на те 14 элементов, которые преобладают и в земной коре, составляя в ней 96,9%, хотя и в других соотношениях. Таким образом, жизнь - это химическое производное литосферы. В организмах обнаружены почти все элементы таблицы Д.И. Менделеева, т.е. они характеризуются той же химией, что и неживая природа.
Биохимические процессы, осуществляющиеся в организмах, представляют собой сложные, организованные в циклы последовательности реакций. На воспроизведение их в неживой природе потребовались бы огромные энергетические затраты. В живых организмах они протекают при посредстве белковых катализаторов - ферментов, понижающих энергию активации реакций на несколько порядков величин. Материалы и энергию для обменных реакций живые существа черпают в окружающей среде. Они преобразуют среду уже только тем, что живут.
Все живое вещество по своей массе занимает ничтожную долю по сравнению с любой из верхних оболочек земного шара. По современным вероятностным оценкам общее количество массы живого вещества в современную эпоху составляет порядка 2420 млрд. т. (Это одна десятимиллионная часть массы литосферы). Однако в качественном отношении живое вещество представляет собой наиболее высокоорганизованную часть материи Земли и наиболее активную форму материи. Оно производит гигантскую геохимическую работу в биосфере, полностью преобразив верхние оболочки Земли за время своего существования.
Геохимическая работа живого вещества
.
Основная планетарная функция живого вещества заключается в создании органического вещества в ходе фотосинтеза, т.е. в связывании и запасании (порой на очень длительное время) солнечной энергии, которая затем идет на поддержание множества других геохимических процессов в биосфере. За время существования жизни на Земле живое вещество превратило в потенциальную энергию органических соединений огромное количество солнечной энергии; значительная часть ее в ходе геологической истории накопилась в связанном виде. Для современной биосферы характерны залежи угля и других органических веществ, образовавшихся в палеозое, мезозое и кайнозое.
В биосфере в результате жизнедеятельности организмов в больших масштабах осуществляются такие химические процессы, как окисление и восстановление элементов с переменной валентностью (азот, сера, железо, марганец и др.). Микроорганизмы-восстановители гетеротрофны, используют в качестве источника энергии органические вещества. К ним относятся денитрифицирующие и сульфатредуцирующие бактерии, восстанавливающие из окисленных форм азот до элементарного состояния и серу до сероводорода. Микроорганизмы-окислители могут быть как автотрофами, так и гетеротрофами. Это бактерии, окисляющие сероводород и серу, нитри- и нитрофицирующие микроорганизмы, железные и марганцевые бактерии, концентрирующие эти металлы в своих клетках.
Геологические результаты деятельности этих организмов проявляются в образовании осадочных месторождений серы, образовании в анаэробных условиях залежей сульфидов металлов, а в аэробных - окисление их и перевод в растворимое состояние, возникновение железных и железомарганцевых руд.
За счет жизнедеятельности огромного числа гетеротрофов, в основном грибов, животных и микроорганизмов, происходит гигантская в масштабах всей Земли, работа по разложению органических остатков. При деструкции органической массы протекают два параллельных процесса. Разложение органических соединений в конечном счете до углекислого газа, аммиака и воды, а в анаэробных условиях еще и до водорода и углеводородов представляет процесс минерализации. Продукты минерализации вновь используются автотрофами. Кроме того, в почве часть освобождающихся веществ ароматической природы под влиянием жизнедеятельности микроорганизмов вновь конденсируется с образованием сложного комплекса соединений - почвенного гумуса (различные гумусовые кислоты и их соли) со значительным запасом энергии. Этот процесс стимулируется деятельностью многих почвенных групп гетеротрофов. Гумус является основой почвенного плодородия. Его разложение протекает в дальнейшем очень медленно, под влиянием определенной, автохтонной микрофлоры почв, чем достигается постоянство в обеспечении растений элементами минерального питания.
Природные воды, обогащенные этими продуктами минерализации, становятся химически высокоактивными и выветривают горные породы.
Процесс разложения органических веществ, при котором освобождается химическая энергия, характерен для всех частей биосферы, где есть живые организмы, тогда как фотосинтез протекает только на поверхности суши и в верхнем слое водоемов. Часть органического вещества, попадающего в условия, неблагоприятные для деятельности деструкторов, захоранивается и консервируется в составе осадочных пород, поэтому синтез органических веществ в масштабе всей биосферы не полностью уравновешивается их разложением.
Эта некоторая несбалансированность процессов синтеза и разложения органических веществ в биосфере определила кислородный режим современной воздушной оболочки Земли.
Кислород атмосферы накоплен за счет фотосинтеза. Единственный источник абиогенного поступления свободного кислорода - фотодиссоциация молекул воды в верхних слоях атмосферы - очень незначителен. Количество молекул О2
, выделяемых зелеными растениями, пропорционально количеству связываемых молекул СО2
. Выделенный кислород вновь используется на окисление углерода при минерализации органического вещества и дыхании организмов, но так как часть органических веществ захоранивается в осадочных породах, то эквивалентное количество О2
остается в атмосфере. Значительная часть его идет на окисление минеральных веществ. С увеличением концентрации свободного кислорода в воздухе расход его на окисление минералов возрастает, с уменьшением - снижается.
В верхних слоях тропосферы под влиянием ультрафиолетового излучения из кислорода образуется озон. Существование озонового экрана также результат деятельности живого вещества, которое, по выражению Вернадского, "как бы само создает себе область жизни".
Углекислый газ поступает в атмосферу за счет дыхания всех организмов. Второй, менее мощный его источник - выделение по трещинам земной коры из осадочных пород за счет химических процессов, совершающихся под действием высоких температур; он также имеет биогенное происхождение. Часть углекислого газа поступает в атмосферу из абиогенного источника - непосредственно из мантии Земли при вулканических извержениях. Эта часть - лишь 0,01% от СО2
, выделенного живыми организмами. Расходуется углекислый газ в процессах органического синтеза, а также на выветривание горных пород и образование карбонатов. Содержание СО2
в атмосфере в ранний период развития жизни было более высоким. В течение фанерозоя оно изменялось в достаточно широком диапазоне. В девоне и начале карбона, а также в перми, по новейшим подсчетам, оно превышало современный уровень в 6-10 раз, а начиная с середины мела неуклонно падает.
Азот атмосферы химически инертен, но и он участвует в процессах синтеза и распада органического вещества. Азот усваивают из атмосферы многие прокариотические организмы - азотфиксаторы; после их смерти он переходит в доступные растениям соединения и включается в цепи питания и разложения.
К газам органического происхождения относятся также сероводород, метан и множество других летучих соединений, создаваемых живым веществом. Продуцируя и потребляя газообразные вещества, организмы биосферы поддерживают постоянство состава воздушной оболочки Земли.
Живое вещество перераспределяет атомы в биосфере. Многие организмы обладают способностью накапливать, концентрировать в себе определенные элементы, несмотря на часто ничтожное содержание их в окружающей среде, например литотамниевые водоросли накапливают в своих телах до 10% магния, в раковинах брахиопод содержится около 20% фосфора, в серных бактериях - до 10% серы. Многие организмы концентрируют кальций, кремний, натрий, алюминий, йод и т.д.; отмирая и захораниваясь в массе, они образуют скопления этих веществ. Возникают залежи таких соединений, как известняки, бокситы, фосфориты, осадочная железная руда и др.
Живое вещество активно участвует также в грандиозных процессах перемещения, миграции атомов в биосфере через систему больших и малых биогеохимических циклов.
Лекция 8.
Биогеохимические циклы.
1. Структура и основные типы биогеохимических циклов.
2. Количественное изучение биогеохимических циклов.
Структура и основные типы биогеохимических циклов.
Подробное рассмотрение биосферы было необходимо не только для осознания космической роли живого вещества в формировании современного облика планеты, оно важно для понимания того, что в отличие от неисчерпаемого потока солнечной энергии, поток вещества для построения всего живого ограничен пределами биосферы. Ввиду того, что растения и животные могут использовать только те биогенные элементы, которые находятся на поверхности Земли или вблизи нее, для сохранения жизни необходимо, чтобы материалы, ассимилированные какими-либо организмами, в конечном счете становились доступными другим организмам. Единственный механизм, способный обеспечить подобную преемственность - это механизм циркуляции элементов протоплазмы из внешней среды в организмы и опять во внешнюю среду. То есть биогенные элементы в отличие от энергии должны удерживаться в экосистеме, где они совершают непрерывный круговорот, в котором участвуют как живые организмы, так и физическая среда.
Известно, что из 90 с лишним элементов, встречающихся в природе 30¸40 требуются живым организмам; некоторые элементы, такие, как углерод, водород, кислород и азот необходимы организмам в больших количествах, другие - в малых или даже в ничтожных количествах.
Какова бы ни была потребность в них, жизненно важные для организмов элементы участвуют в круговоротах. Элементы, не имеющие столь существенного значения, хотя и не так тесно связаны с организмами, но также участвуют в циклическом движении. Часто они движутся теми же путями, что и незаменимые элементы, из-за своего химического родства с последними.
Эти в большей или меньшей степени замкнутые пути получили название биогеохимических циклов. Название четко отражает суть того, что обмен веществ осуществляется между живыми и неживыми компонентами биосферы.
В природе элементы никогда, или почти никогда не распределены равномерно по всей экосистеме и не находятся всюду в одной и той же химической форме.
Например, кислород содержится в атмосфере в газообразной форме - в виде молекулярного кислорода (О2
) и в виде диоксида углерода (СО2
); в воде кислород содержится в растворенном виде, но, кроме того, он в соединении с водородом входит в состав самой воды (Н2
О). В литосфере кислород встречается в форме оксидов (Fе2
О3
и др.) и солей (главным образом в виде CаСО3
). Скорость перехода элемента из одного неорганического соединения в другое и его доступность в неорганической форме живым организмам сильно варьируют. Самый большой фонд кислорода, в который входит свыше 90% всего кислорода, находящегося у поверхности Земли, - это карбонат кальция осадочных пород, в частности известняков; за исключением небольших количеств, освобождаемых в результате вулканической деятельности, кислород, входящий в состав известняков и других осадочных пород, совершенно недоступен живым организмам. В отличие от кислорода азот встречается, главным образом, в газообразной форме (N2
) в атмосфере, однако растения ассимилируют азот в основном из нитратов (NO3-
), содержащихся в почве или в воде. Несмотря на свое обилие, атмосферный азот играет незначительную роль в круговороте питательных веществ.
Таким образом, в каждом круговороте удобно различать две части, или два "фонда":
1) Резервный фонд - большая масса медленно движущихся веществ, в основном небиологический компонент, то есть та часть круговорота, которая физически или химически отделена от организмов;
2) Подвижный (или обменный фонд) - меньший, но более активный, для которого характерен быстрый обмен между организмами и их непосредственным окружением.
Иногда резервный фонд называют "недоступным" фондом, а активный, циркулирующий фонд - доступным или обменным. Такие термины допустимы, если только не понимать их слишком буквально. Любой атом, находящийся в резервном фонде не обязательно все время недоступен для организмов, так как между доступным и недоступным фондами существует постоянный медленный обмен.
Экосистему удобно представить в виде ряда блоков, через которые проходят различные материалы и в которых эти материалы могут оставаться на протяжении различных периодов времени (рис.6).
В круговоротах минеральных веществ в экосистеме в большинстве случаев участвуют три активных блока: живые организмы, мертвый органический детрит и доступные неорганические вещества. Два добавочных (резервных) блока - косвенно доступные неорганические вещества и осаждающиеся органические вещества - связаны с круговоротами биогенных элементов в каких-либо периферических участках, однако обмен между этими блоками и остальной экосистемой замедлен по сравнению с обменом, происходящим между активными блоками.
Процессы, обеспечивающие перенос биогенных элементов в пределах экосистемы, представлены на рис.6. Ассимиляция и создание продукции сопровождаются переходом минеральных веществ из неорганического блока в органический: в круговороте углерода, кислорода, азота, фосфора и серы самым главным компонентом этого этапа является первичная продукция, создаваемая растениями; однако животным необходимы, кроме того, многие другие важные элементы, такие, как натрий, калий и кальций, и они ассимилируют эти элементы непосредственно из воды, которую пьют.
Некоторая часть углерода и кислорода возвращается в результате дыхания непосредственно в фонд доступных неорганических питательных веществ, возможно, после многократных круговоротов в пределах блока живой биомассы по хищным пищевым цепям.
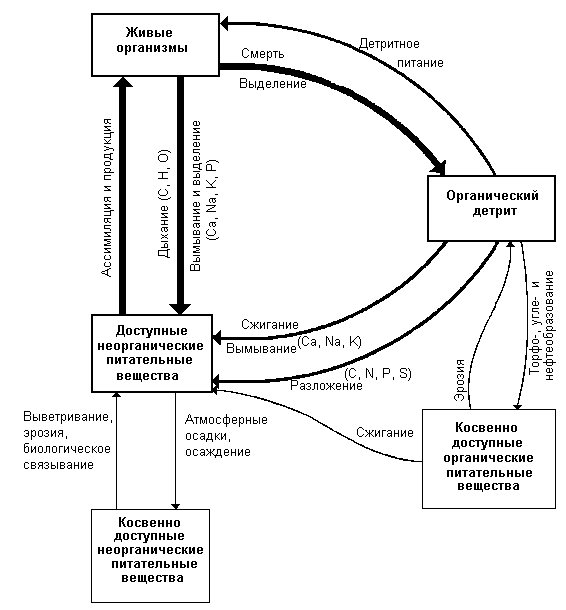
Рис.6. Блочная модель экосистемы с указанием некоторых наиболее важных путей обмена минеральных веществ.
Кальций, натрий и ионы других минеральных веществ выделяются или вымываются из листьев дождем или водой, окружающей водные организмы, и тоже быстро вновь вступают в круговорот. Большая часть углерода и азота, включившихся в процессе ассимиляции в живую биомассу, после гибели организмов, а также в результате экскреции переносится в детритный блок. Некоторые биогенные элементы, содержащиеся в детрите, могут быть возвращены в блок биомассы детритоядными организмами, но все они в конечном счете в результате вымывания и разложения вновь попадают в фонд доступных неорганических веществ. Обмен между фондами активно участвующих в круговороте минеральных веществ и огромными резервуарами косвенно доступных биогенных элементов, заключенных в атмосфере, известняках, каменном угле и в образующих земную кору горных породах, происходит медленно, главным образом, в результате геологических процессов.
Процессы ассимиляции и распада, благодаря которым происходят круговороты биогенных элементов в биосфере, тесно связаны с поглощением и высвобождением энергии организмами. Наиболее тесно связан с превращениями энергии в сообществе круговорот углерода, так как большая часть энергии, ассимилированной в процессе фотосинтеза, содержится в органических углеродсодержащих соединениях. В большинстве процессов, сопровождающихся выделением энергии, среди которых самым главным является дыхание, углерод высвобождается в виде диоксида углерода. Когда в организме происходит метаболизм органических соединений, содержащих азот, фосфор и серу, эти элементы нередко удерживаются в нем, поскольку они необходимы для синтеза структурных белков, ферментов и других органических молекул, образующих структурные и функциональные компоненты живых тканей. Поэтому прохождение азота, фосфора и серы через каждый трофический уровень несколько замедленно по сравнению со средним временем переноса энергии.
При оценке влияния деятельности человека на биогеохимические циклы важны сравнительные объемы резервных фондов. Как правило, изменениям подвержены, в первую очередь, наиболее малообъемные и малоподвижные фонды.
Разделение биогеохимических циклов на круговороты газообразных веществ и осадочные циклы основано на том, что некоторые круговороты, например, те, в которых участвуют углерод, азот или кислород, благодаря наличию крупных атмосферных или океанических (или же и тех и других) фондов довольно быстро компенсируют различные нарушения. Например, избыток СО2
, накопившийся в каком-либо месте в связи с усиленным окислением или горением, обычно быстро рассеивается воздушными потоками; кроме того, усиленное образование СО2
компенсируется усиленным его потреблением растениями и превращением в карбонаты в море. Круговороты газообразных веществ с их большими атмосферными фондами можно считать в глобальном масштабе "хорошо забуференными", поскольку их способность приспосабливаться к изменением велика. Но способность к саморегуляции даже при таком большом резервном фонде, каким является атмосфера, имеет свои пределы. Осадочные циклы, в которых участвуют такие элементы, как фосфор и железо, обычно гораздо менее подвержены самоконтролю и легче нарушаются в результате местных пертурбаций, потому, что в этих случаях основная масса вещества сосредоточена в относительно малоактивном и малоподвижном резервном фонде в земной коре. Следовательно, если "спуск" совершается быстрее, чем обратный "подъем", то какая-то часть обменного материала на длительное время выбывает из круговорота; механизмы, обеспечивающие возвращение в круговорот, во многих случаях основаны, главным образом, на биологических процессах.
Человек уникален не только тем, что в своей деятельности он использует почти все имеющиеся в природе элементы, а также ряд новых, искусственно им созданных. Он так ускоряет движение многих веществ, что круговороты становятся несовершенными или процесс теряет цикличность и складывается противоестественная ситуация: в одних местах возникает недостаток, а в других - избыток каких-то веществ. В этой связи первоочередная задача - количественное изучение биогеохимических циклов.
Количественное изучение биогеохимических циклов
Количественное изучение биогеохимических циклов включает изучение скоростей циркуляции и оценку имеющихся запасов.
Знание объемов фондов необходимо для расчета возможных нагрузок на экосистему.
Для определения же структуры и функции экосистемы важнее оценить скорости обмена или переноса веществ, нежели количество веществ, находящихся в данное время в данном месте. Действительно, низкое содержание доступного вещества может означать, что система либо истощена, либо ее метаболизм весьма интенсивен; понять ситуацию, оценить продуктивность системы можно только в том случае, если измерить скорость потока элемента. Для высокопродуктивных систем и для поддержания высокого уровня продукции органических веществ скорость миграции важнее, чем концентрация элемента.
Очень важный для практики вывод, вытекающий из многих интенсивных исследований круговоротов биогенных элементов, состоит в том, что избыток удобрений может оказаться столь же невыгодным для человека, как и их недостаток; если в систему вносится больше веществ, чем могут использовать активные в данный момент организмы, излишек быстро связывается почвой и отложениями или исчезает в результате выщелачивания, становясь недоступным именно в тот период, когда рост организмов наиболее желателен. Более того, неразумные субсидии могут обернуться источником стресса.
Для сравнения скоростей обмена между разными компонентами экосистемы вводят понятие оборота. Если говорить об обмене после установления равновесия, то скорость оборота - это та часть общего количества данного вещества в данном компоненте системы, которая высвобождается (или поглощается) за определенное время; а время оборота представляет собой обратную величину, то есть время необходимое для полной смены всего количества этого вещества в данном компоненте экосистемы. Например, если в компоненте содержится 1000 ед. вещества и в 1 час поступает или убывает 10 ед., то скорость оборота равна 10/1000, или 0,01, т.е. 1% в час. Время оборота будет равно 1000/10, или 100 часам.
Количественные характеристики биогеохимических циклов изучены еще недостаточно, особенно в крупных системах. В последние 30 - 40 лет с усовершенствованием разнообразных, используемых в экологии современных методов, в том числе метода радиоактивных индикаторов, масс-спектрометрии, автоматического слежения и дистанционных измерений, появилась возможность измерять скорости циркуляции в довольно обширных экосистемах (озеро, лес) и приступить к самой важной задаче - получить количественную оценку биогеохимических круговоротов в глобальном масштабе.
Применение радионуклидов в качестве индикаторов, или "меток" дало огромный толчок этим исследованиям. В эксперименте в экосистему или в отдельные организмы вводят изотоп в крайне малых количествах по сравнению с уже имеющимися в системе количествами нерадиоактивного элемента, так что в системе не происходит никаких нарушений ни за счет радиоактивности, ни за счет изменения концентрации. Все, что происходит с меткой (даже самые малые количества которой выявляются благодаря ее заметному излучению), отражает то, что обычно происходит в системе с интересующим нас элементом. Использование радионуклидов также открыло возможность анализа разных частей экосистемы посредством составления блоковых схем и разработки и "настраивания" усовершенствованных математических моделей.
Наиболее интересный вывод, к которому приводит обобщение и анализ имеющихся результатов по количественным оценкам биогеохимических циклов заключается в том, что вид не обязательно должен входить в пищевую цепь человека, чтобы быть полезным ему. Многие виды приносят пользу косвенным путем, что остается незамеченным при поверхностном взгляде. Как показывают сейчас наблюдения и усовершенствованные модели, причинно-следственные взаимоотношения между многими видами управляют переносом веществ и энергии в компоненты экосистем, которые непосредственно связаны о человеком.
Лекция 9.
Круговороты основных биогенных элементов.
1. Глобальный круговорот воды.
2. Глобальный круговорот углерода.
Глобальный круговорот воды
Круговороты воды и СО2
в глобальном масштабе представляют собой, вероятно, самые важные для человечества биогеохимические круговороты. Для обоих характерны небольшие, но весьма подвижные фонды в атмосфере, высокочувствительные с нарушениям, которые вызываются деятельностью человека и которые могут влиять на погоду и климат.
Несмотря на то, что вода участвует в химических реакциях, из которых слагается и фотосинтез, большая часть потока воды, проходящего через экосистему, связана с испарением, транспирацией (испарение растениями) и выпадением осадков.
Круговорот воды, или гидрологический цикл, как и любой другой круговорот, приводится в движение энергией. Поглощение световой энергии жидкой водой представляет собой главную точку, в которой источник энергии сопряжен с круговоротом воды. По оценкам, около трети всей поступающей на Землю солнечной энергии затрачивается на приведение в движение круговорота воды.
Более 90 % имеющейся на земном шаре воды связано в горных породах, образующих земную кору, и в отложениях (льда и снега) на поверхности Земли. Эта вода вступает в происходящий в экосистеме гидрологический цикл очень редко: лишь при вулканических выбросах водяных паров. Таким образом, большие запасы воды, имеющиеся в земной коре, вносят весьма незначительный вклад в передвижение воды вблизи поверхности Земли, составляя основу резервного фонда этого круговорота.
Фонд воды в атмосфере невелик (составляет около 3%). Вода, содержащаяся в воздухе в виде пара в любой данный момент, соответствует в среднем слою толщиной 2,5 см, равномерно распределенному по поверхности Земли. Количество осадков, выпадающих за год, составляет в среднем 65 см, что в 25 раз больше того количества влаги, которое содержится в атмосфере в любой данный момент. Следовательно, водяные пары, постоянно содержащиеся в атмосфере, так называемый атмосферный фонд, ежегодно совершают круговорот 25 раз. Соответственно время переноса воды в атмосфере равно в среднем двум неделям.
Содержание воды в почве, реках, озерах и океанах в сотни тысяч раз больше, чем в атмосфере. Однако она протекает через оба эти фонда с одинаковой скоростью, поскольку испарение сбалансировано с выпадением осадков. Среднее время переноса воды в ее жидкой фазе по поверхности Земли, равное 3650 годам, в 105
раз больше, чем время ее переноса в атмосфере.
Особое внимание следует обратить на следующие аспекты круговорота воды:
1. Море теряет из-за испарения больше воды, чем получает с осадками; на суше ситуация противоположная. Т.о. значительная часть осадков, поддерживающих экосистемы суши, в том числе большинство агроэкосистем, состоит из воды, испаренной из моря.
2. Важная, если не главная роль транспирации
растений в общей эвапотранспирации (испарении) с суши. Влияние, оказываемое растительностью на движение воды, выявляется лучше всего при удалении растительности. Так экспериментальная вырубка всех деревьев в бассейнах небольших рек увеличивает сток воды в реки, дренирующие расчищенные участки, более чем на 200%. В нормальных условиях этот излишек в виде водяного пара траспирировался бы непосредственно в атмосферу.
3. Хотя поверхностный сток пополняет резервуары грунтовых вод и сам пополняется от них, эти величины имеют обратную зависимость. В результате деятельности человека (покрытия земной поверхности непроницаемыми для воды материалами, создания водохранилищ на реках, строительства оросительных систем, уплотнения пахотных земель, сведения лесов и т.д.) сток увеличивается и пополнение столь важного фонда грунтовых вод сокращается. Во многих засушливых районах резервуары грунтовых вод сейчас быстрее выкачиваются человеком, чем
пополняются природой.
Глобальный круговорот углерода.
В биологическом круговороте углерода участвуют только органические соединения и диоксид углерода; фотосинтез и дыхание полностью комплементарны. Весь ассимилированный в процессе фотосинтеза углерод включается в углеводы, а в процессе дыхания углерод, содержащийся в органических соединениях, превращается в диоксид углерода.
Обширные фонды углерода неорганического происхождения - атмосферный диоксид углерода, растворенный диоксид углерода (главным образом в форме HCO3
-
), угольная кислота и карбонатные отложения - участвуют в круговороте углерода в различной степени . Обмен между углеродом, содержащимся
в изверженных
породах, отложениях карбоната кальция, каменном угле и
нефти, и другими более активными его фондами происходит настолько медленно, что влияние этого углерода на краткосрочное функционирование экосистем незначительно.
В круговороте СО2
атмосферный фонд очень невелик, в сравнении с запасами углерода в океанах, в ископаемом топливе и других резервуарах земной коры. Полагают, что до наступления индустриальной эры потоки углерода между атмосферой, материками и океанами были сбалансированы.
В основе этого баланса лежит регулирующая деятельность зеленых растений и поглощающая способность карбонатной системы моря. Когда более 2 млрд. лет назад на Земле появилась жизнь, атмосфера состояла из вулканических газов. В ней было много СО2
и мало кислорода (а быть может, его не было совсем), и первые организмы были анаэробными. В результате того, что продукция в среднем слегка превосходила дыхание, за геологическое время в атмосфере накопился кислород и уменьшилось содержание СО2
. Накоплению кислорода способствовали также геологические и чисто химические процессы, например высвобождение его из оксидов железа или образование восстановленных соединений азота и расщепление воды ультрафиолетовым излучением с выделением кислорода. Низкое содержание СО2
, также как высокие концентрации О2
служат лимитирующими факторами для фотосинтеза: для большинства растений характерно увеличение интенсивности фотосинтеза, если в эксперименте увеличивается содержание CО2
или понижается содержание О2
. Таким образом, зеленые растения оказываются весьма чувствительным регулятором содержания этих газов.
Фотосинтезирующий "зеленый пояс" Земли и карбонатная система моря поддерживают постоянный уровень содержания СО2
в атмосфере. Но в последнем столетии стремительно возрастающее потребление горючих ископаемых вместе с уменьшением поглотительной способности "зеленого пояса" начинает превосходить возможности природного контроля, так что содержание СО2
в атмосфере, сейчас постепенно возрастает. Действительно, наибольшим изменениям подвержены потоки веществ на входе и на выходе небольших обменных фондов. Полагают, что
в начале промышленной революции (примерно 1800 г.) в атмосфере Земли содержалось около 290 частей СО2
на миллион
(0,029 %). В 1958 г., когда были впервые проведены точные измерения, содержание составило 315, а в 1960 г. оно выросло до 335 частей на миллион. Если концентрация в
двое превысит доиндустриальный
уровень, что может случишься
к середине будущего века, вероятно потепление климата Земли: температура в среднем повысится на 1,5 - 4,5°С,
и это наряду с подъемом уровня моря (в результате таяния полярных шапок) и изменением распределения осадков может погубить сельское хозяйство.
Считают, что в следующем веке может установиться новое, но ненадежное равновесие между увеличением содержания СО2
(способствующего разогреву Земли) и усилением загрязнения атмосферы
пылью и другими частицами, отражающими излучение и этим охлаждающими планету. Любое
значительное результирующее изменение теплового бюджета
Земли тогда повлияет на климат.
Основным
источником поступления "парникового газа" СО2
считается сжигание горючих ископаемых, однако свой вклад вносят также развитие сельского хозяйства и сведение лесов. Может показаться удивительным, что сельское хозяйство в конечном счете приводит к потере СО2
из почвы (то есть вносит в атмосферу больше, чем забирает оттуда), но дело в том, что фиксация СО2
сельскохозяйственными культурами, многие из которых активны лишь часть года, не компенсирует количества СО2,
высвобождающееся из почвы, особенно в результате частой вспашки. Леса
- важные накопители углерода, так как в биомассе лесов содержится в 1,5 раза, а в лесном гумусе - в 4 раза больше углерода, чем в атмосфере. Сведение леса, разумеется, может высвободить углерод, накопленный в древесине, особенно если она немедленно сжигается. Уничтожение леса, особенно при последующем использовании этих земель для сельского хозяйства или строительства городов, приводит к окислению
гумуса.
Кроме СО2
в атмосфере присутствуют в небольших количествах еще два соединения углерода: оксид углерода (СО)- примерно 0,1 части на миллион и метан (СH4
) - около 1,6 части на миллион. Как и СО2
, эти соединения находятся в быстром круговороте и поэтому имеют небольшое время пребывания в атмосфере - около 0,1 года для СО; 3,6 года для СH4
и 4 года для СО2
.
И СО,
и СH4
образуются при неполном или анаэробном разложении органического вещества; в атмосфере оба окисляются до СО2
. Столько же СО , сколько попадает в атмосферу в результате естественного разложения, вносится в нее сейчас при неполном сгорании горючих ископаемых, особенно с выхлопными газами.
Накопление монооксида углерода - этого смертельного яда для человека - в глобальном масштабе не представляет собой угрозы, но в городах, где воздух застаивается, повышение концентрации этого газа в атмосфере начинает становиться угрожающим, достигая 100 частей на миллион.
Производство метана - одна из важнейших функций водно-болотистых угодий и мелководных морей мира. Метан, как полагают, имеет полезную функцию: он поддерживает стабильность озонного слоя в верхней атмосфере, который блокирует смертельно опасное ультрафиолетовое излучение Солнца.
Лекция 10.
Круговороты основных биогенных элементов.
1. Круговорот кислорода.
2. Типы фотосинтеза и организмов-продуцентов.
3. Типы катаболизма и организмов-разрушителей.
4. Общий баланс процессов продукции и разложения.
Круговорот кислорода .
Вторым по содержанию в атмосфере после азота является кислород, составляющий 20,95% ее по объему. Гораздо большее его количество находится в связанном состоянии в молекулах воды, в солях, а также в оксидах и других твердых породах земной коры, однако к этому огромному фонду кислорода экосистема не имеет непосредственного доступа. Время переноса кислорода в атмосфере составляет около 2500 лет, если пренебречь обменом кислорода между атмосферой и поверхностными водами.
Механизм круговорота кислорода достаточно прост. Полагают, что молекула кислорода (О2
) , образующаяся при фотосинтезе, получает один свой атом от диоксида углерода, а другой - от воды; молекула кислорода, потребляемая при дыхании, отдает один свой атом диоксиду углерода, а другой - воде. Таким образом, круговорот кислорода завязан на процессы фотосинтеза и дыхания, с которыми интересно будет познакомиться.
Типы фотосинтеза
и организмов-продуцентов.
С химической точки зрения процесс фотосинтеза включает запасание
части энергии
солнечного света в виде
потенциальной энергии пищи. Общее уравнение окислительно-восстановительной реакции можно записать следующим образом:
СО2
+2 H2
A « {СН2
O} + H2
О +2A
окисление описывается уравнением:
H2
A - 2e-
® 2H+
+A;
а восстановление :
СО2
+4H+
+ 4e-
® {СН2
O}+ H2
О.
Для зеленых растений (водорослей, высших растений) А - это кислород; вода окисляется с высвобождением газообразного кислорода, а диоксид углерода восстанавливается до углеводов ({СН2
O}) с высвобождением воды. Такой тип фотосинтеза носит название "нормального фотосинтеза".
При бактериальном фотосинтезе, напротив, H2
A
- восстановитель - не вода, а либо неорганическое соединение серы, например сероводород H2
S, как у зеленых и пурпурных серобактерий, либо органическое соединение, как у пурпурных и бурых несерных
бактерий. Соответственно при бактериальном фотосинтезе этих типов кислород не выделяется.
Фотосинтезирующие
бактерии в основном водные морские и пресноводные организмы; в большинстве случаев они играют незначительную роль в продукции органического вещества. Но они способны функционировать в условиях, в общем неблагоприятных для большинства зеленых растений, и в водных отложениях участвуют в круговороте некоторых элементов. Бактериальный фотосинтез
может быть полезен в загрязненных и эвтрофных
(кормных)
водах, восстановленных зонах
с ограниченным доступом света,
в стоячих озерах, богатых сероводородом, где на его долю может приходиться до 30% общей продукции.
Установлено,
что у высших растений существуют разные биохимические механизмы восстановления диоксида углерода до углеводов, сопровождающегося
выделением кислорода. У большинства растений фиксация диоксида углерода идет по С3
- пентозофосфатному
пути, или циклу Кальвина.
Иной путь - по циклу С4
- дикарбоновых
кислот. При обсуждении экологических последствий этих особенностей растения в соответствии с характером протекающих у них процессов фотосинтеза называют С3
-растениями или С4
-
растениями. Эти растения по-разному реагируют на свет, температуру и воду. У первых максимальная интенсивность фотосинтеза
(на единицу поверхности листа) обычно наблюдается при умеренных освещенности и температуре, а высокие температуры и яркий солнечный свет подавляют фотосинтез.
Напротив, С4
-
растения адаптированы к яркому свету и высокой температуре и в таких условиях значительно превосходят по продуктивности С3
-растения.
Кроме того,
они более эффективно используют воду: как правило, на производство 1 г
сухого вещества им требуется менее 400 г
воды, а С3
- -
растениям - от 400 до
1000 г
воды. К тому же фотосинтез у С4
-
растений не ингибируется
высокими концентрациями кислорода, как это происходит у С3
-
видов. Одна из причин того, что С4
-
растения более эффективны у верхних пределов световой
и температурной шкал, состоит в том, что у них невелико фотодыхание,
т.е. при увеличении освещенности продукты фотосинтеза
не тратятся на дыхание.
Хотя в пересчете на площадь листвы эффективность фотосинтеза у С3
-
растений ниже, эти растения создают большую
часть фотосинтетической
продукции мира,
возможно потому, что они более конкурентноспособны в смешанных сообществах,
где растения затеняют друг друга и где освещенность, температура и другие показатели
ближе
к средним значениям, чем к предельным. Это еще о
дин хороший пример принципа
эмерджентности.
Выживание наиболее приспособленных в реальном мире - не всегда выживание видов, физиологически более эффективных
в оптимальных условиях
в монокультуре; чаще выживают виды, преуспевающие в смешанной культуре в изменчивых и не всегда оптимальных условиях. Иными словами, то, что эффективно в изоляции, не
обязательно эффективно
в сообществе, где на естественный отбор сильно влияют межвидовые взаимодействия.
Как и следовало ожидать, С4
-
виды преобладают среди растительности пустынь и степей в теплом и тропическом климате и редки в лесах и на севере, где освещенность и температура низкие.
Недавно был открыт еще один способ фотосинтеза, приспособленный к условиям пустынь и получивший название САМ - метаболизм (кислотный метаболизм толстенковых).
У некоторых жителей пустыни, в том числе кактусов, устьица на протяжении жаркого дня закрыты и открываются только прохладной ночью. Диоксид углерода, поглощаемый через устьица, накапливается в форме органических кислот и фиксируется в углеводах только на следующий день. Такая задержка фотосинтеза
значительно уменьшает дневные потери воды, усиливая этим способность растений сохранять водный баланс и запасы воды.
Микроорганизмы, которых называют хемосинтезирующими
бактериями, относят к хемолитотрофам,
потому что они получают энергию для включения диоксида углерода в состав компонентов клетки не за счет фотосинтеза, а в результате химического окисления простых неорганических соединений, например аммиака (в нитрит), нитрита (в нитрат), сульфида (в серу), двухвалентного железа (в трехвалентное). Такие микроорганизмы могут расти в темноте, но большинству из них нужен кислород.
В качестве примера можно привести различные азотные бактерии, играющие
важную роль в круговороте а
зота. Благодаря способности функционирования в отсутствие света - в осадках, почве и на дне океанов- хемосинтезирующие
бактерии не только играют роль в извлечении минеральных питательных веществ, они используют энергию, которая иначе была бы недоступна консументам.
Большинство высших (семенных) растений и многие виды
водорослей используют
только простые неорганические вещества и, следовательно, являются полностью автотрофными.
Но некоторые водоросли нуждаются в каком-то одном (
определенном) сложном органическом "ростовом веществе", которое они сами не способны синтезировать. Другие виды нуждаются в двух, трех или многих таких "ростовых веществах" и, следовательно, являются частично гетеротрофными; организмы, занимающие промежуточное положение между автотрофами
и гетеротрофами, называются ауксотрофными.
В глобальном масштабе эволюционно
наиболее развитые формы жизни можно четко разделить на автотрофов
и гетеротрофов,
причем для выживания последних необходим газообразный кислород. Но многие виды и штаммы низших микроорганизмов
- бактерий, грибов, низших водорослей и простейших - не столь специализированы, они приспособлены к промежуточному способу существования и могут с автотрофии
переключаться на гетеротрофию,
жить в присутствии и в отсутствии кислорода.
Типы катаболизма и организмов-разрушителей.
Катаболизм (разложение) органических остатков - длительный и сложный процесс, контролирующий
несколько важных функций экосистемы. В
результате этого
процесса:
1) возвращаются в
круговорот элементы питания, находящиеся
в мертвом органическом
веществе;
2) производится пища для последовательного ряда организмов в детритной
пищевой цепи;
3) производятся вторичные метаболиты ингибирующего,
стимулирующего и часто регулирующего действия;
4) образуются хелатные комплексы с элементами питания;
5) преобразуются инертные вещества земной
поверхности, что приводит к образованию такого уникального природного тела, каким является почва;
6) поддерживается состав атмосферы, способствующий жизни крупных аэробов,
таких, как человек.
Если
рассматривать разложение в широком смысле слова, как "любое биологическое окисление, дающее энергию", то с учетом потребности в кислороде можно выделить несколько типов этого
процесса, приблизительно аналогичных типам фотосинтеза:
1. Аэробное дыхание - окислителем (акцептором
электронов) служит газообразный молекулярный кислород (тип 1 );
2. Анаэробное дыхание протекает без участия газообразного кислорода. Акцептором электронов служит не кислород, а какое-либо другое неорганическое (тип 2) или органическое (тип 3) соединение;
3. Брожение тоже анаэробный процесс, но окисляемое органическое соединение само служит акцептором
электронов (тип 4).
Аэробное дыхание (тип 1) - процесс обратный "нормальному фотосинтезу"; в этом процессе синтезированное органическое вещество {СН2
O} вновь разлагается с образованием СО2
и H2
О и с высвобождением энергии. Все высшие растения и животные и большинство микроорганизмов получают энергию для поддержания жизнедеятельности и построения клеток именно с помощью этого процесса. В итоге завершенного дыхания образуются СО2
,
вода и вещества клетки; однако процесс может идти не до конца, и в результате такого незавершенного дыхания образуются органические соединения, еще содержащие некоторое количество энергии, которая в дальнейшем может быть использована другими организмами (процессы 2 и 3).
Бескислородное дыхание служит основой жизнедеятельности
главным образом
у сапрофагов
(бактерии, дрожжи, плесневые грибы, простейшие), хотя, как звено метаболизма, оно может встречаться и
в некоторых тканях высших животных. Хороший пример облигатных
анаэробов - метановые
бактерии, которые разлагают органические соединения, образуя метан
путем восстановления, либо органического углерода, либо углерода карбонатов. Таким образом, дыхание у них может происходить по типам 2 и
3.
К общеизвестным организмам, использующим брожение (тип 4), относятся дрожжи; они
имеют большую практическую ценность для человека, но, кроме того, в изобилии встречаются в почве, где играют ключевую роль в разложении
растительных остатков.
Многие группы бактерий (например факультативные анаэробы) способны и к аэробному и к анаэробному дыханию. Однако конечные продукты этих двух процессов различны, и количество высвобождающейся энергии при анаэробном дыхании значительно меньше.
Общий баланс процессов продукции и разложения
Каждый год фотосинтезирующими
организмами на Земле создается около 100 млрд. т.
органического вещества. За этот промежуток времени приблизительно такое же количество живого вещества окисляется, превращаясь в СО2
и воду в результате дыхания организмов. Однако этот баланс неточен. Для биосферы в целом важнейшее значение
имеет отставание процесса
полной гетеротрофной
утилизации и разложения
продуктов автотрофного
метаболизма от процесса их создания, поскольку именно отставание обусловило накопление в недрах горючих ископаемых, а в атмосфере - кислорода. В этой
связи крайнюю озабоченность вызывает деятельность
человека, который хотя и ненамеренно,
но очень значительно
ускоряет процессы разложения.
Лекция 11.
Круговороты азота и серы.
1. Круговорот азота.
2. Круговорот серы.
Круговорот азота.
Круговорот азота - пример очень сложного и хорошо забуференного круговорота газообразных веществ. Воздух, на 78% состоящий из азота, представляет собой крупнейший "резервуар" и одновременно "предохранительный клапан" системы. Азот постоянно поступает в атмосферу благодаря деятельности денитрифицирующих бактерий и постоянно возвращается в круговорот в результате деятельности азотфиксирующих бактерий или водорослей (биологическая фиксация азота), а также действию физических процессов (например молний), в которых происходит фиксация азота.
Путь прохождения азота через экосистему отличается от пути углерода и кислорода в нескольких важных аспектах. Во-первых, большинство организмов не могут ассимилировать азот из огромного его фонда (3,85·1021
г N2
), имеющегося в атмосфере. Во-вторых, азот не принимает непосредственного участия в высвобождении химической энергии при дыхании: главная его роль сводится к тому, что он входит в состав белков и нуклеиновых кислот, которые создают структуру биологических систем и регулируют их функционирование. В-третьих, биологическое разложение азотсодержащих органических соединений до неорганических форм слагается из нескольких стадий, и некоторые из этих стадий могут осуществляться только специализированными бактериями. В-четвертых, большая часть биохимических превращений, участвующих в разложении азотсодержащих соединений, происходит в почве, где доступность азота растениям облегчается растворимостью его неорганических соединений.
Содержание азота в живых тканях составляет чуть больше 3 % содержания его в активных фондах экосистемы; остальной азот распределен между детритом и нитратами, содержащимися в почве и океане. Кроме того, относительно небольшие количества азота находятся на промежуточных стадиях разложения белка - в виде аммиака и нитритов (табл. 3). Растения ежегодно ассимилируют 86·1014
г азота - менее 1 % активного фонда, поэтому общее время круговорота азота превышает 100 лет.
При круговороте азота происходит поэтапный распад органических соединений, в котором участвует много разных организмов и в результате которого азот в конечном счете переходит в нитратную форму.
Из всех доступных растениям форм, в каких азот содержится в почве, наиболее желательной является аммиак (NН3
) или ион аммония (NН4
+
), потому что их превращение в органические соединения требует минимальных химических перестроек. Аммиак, однако, не может служить источником азота в почве потому, что в высоких концентрациях он токсичен для растительных тканей, и также потому, что он не удерживается в почве. Аммиак легко растворяется в воде и быстро вымывается из почвы. В кислых почвах аммиак превращается в ион аммония. Этот положительно заряженный ион в результате электростатического взаимодействия может присоединяться к поверхности глинисто-гумусовой мицеллы, однако он легко вытесняется в кислых почвах ионами водорода и тем самым тоже довольно легко вымывается водой. Некоторые глинистые минералы просто адсорбируют ионы аммония, включая их в свою кристаллическую решетку, и притом так прочно, что эти ионы не поддаются вымыванию и тем самым становятся недоступными для растений. Тот аммиак, который избежал вымывания из почвы, подвергается действию специализированных бактерий, извлекающих энергию путем окисления азота аммиака до нитритов (NО2
-
) и нитратов (NO3
-
). Отрицательно заряженные нитрит- и нитрат-ионы совершенно не связываются с частицами глины, а поэтому легко вымываются. Образовавшиеся в почве нитраты быстро ассимилируются корнями растений. В наземных экосистемах главные запасы азота представляет азот, входящий в состав органического детрита. В водных экосистемах азот содержится главным образом в виде растворенных нитратов.
Таблица 3.
Распределение азота между активными фондами и годовые скорости переноса
(все фонды содержат в сумме около 1018
г азота )
| Фонд |
Азот, % |
Скорость переноса за год, % |
Органические формы
Растения
Животные
Детрит
Неорганические формы в почвах и океанах
Аммиак (NН4
+
в пересчете на NН3
)
Нитриты (NО2
-
)
Нитраты (NO3
-
)
|
11
11
6100
286
138
4180
|
25
1,4
30
63
2,1
|
Биохимические превращения азотсодержащих соединений чрезвычайно разнообразны, потому что азот может соединяться с другими элементами несколькими различными способами. Наиболее важные процессы в круговороте азота - это распад органических азотсодержащих соединений в результате аммонификации и нитрификации, восстановление нитратов и нитритов до молекулярного азота (N2
) в результате денитрификации и его высвобождение в атмосферу, а также процесс биологической ассимиляции атмосферного азота путем его фиксации.
В процессе денитрификации азот удаляется из активных фондов почвы и поверхностных слоев воды и попадает в атмосферу; в результате же процесса фиксации атмосферный азот возвращается в активный круговорот, происходящий в экосистеме. Эти процессы представляются второстепенными по сравнению с общим круговоротом азота в экосистеме, однако в тех местах, где содержание азота в почве недостаточно для нормального роста растений, фиксация азота нередко приобретает важное значение .
В органических соединениях азот обычно представлен амино- или какой-либо родственной группой, входящей в состав той или иной органической молекулы. У животных выведение из организма избыточного азота происходит путем отщепления аминов от органических соединений и выделения их в сравнительно неизменной форме, главным образом в виде аммиака (NН3
) или мочевины СО(NH2
)2
. Почвенные микроорганизмы легко превращают мочевину в аммиак путем гидролиза:
СО(NH2
)2
+ H2
O ® 2 (NН3
) + CO2
Эта реакция, однако, не сопровождается высвобождением энергии, которая могла бы использоваться для выполнения каких-либо биологических функций.
Некоторые специализированные, и повсеместно встречающиеся бактерии могут высвобождать химическую энергию, содержащуюся в аминогруппе, в результате ряда реакций нитрификации, для которых необходим кислород. Nitrosomonas превращает ион аммония в нитрит; Nitrobacter завершает процесс нитрификации, окисляя нитрит до нитрата.
Нитрификация представляет собой решающий этап в круговороте азота, так как она в конечном счете определяет скорость, с которой азот переходит в форму, доступную зеленым растениям, и тем самым оказывает влияние на продуктивность местообитания. Любые почвенные условия, подавляющие активность бактерий, - высокая кислотность и плохая аэрация почвы, низкая температура и недостаток влаги - подавляют также и нитрификацию. Медленное поступление в почву биогенных элементов, характерное для холодных и засушливых условий, может, в дополнение к непосредственному воздействию этих факторов на фотосинтез, еще больше снижать продуктивность растений. Кроме того, если содержание азота в органическом детрите невелико по сравнению с содержанием в нем углерода, то бактерии расходуют весь этот азот на построение своих клеток, а не используют его в качестве субстрата для метаболизма. В результате азот оказывается связанным в биомассе бактерий, вместо того, чтобы стать доступным растениям.
Денитрификация, в процессе которой нитраты превращаются в азот, происходит в несколько этапов:
NO3
-
® NО2
-
® N2
O ® N2
,
причем на каждом из этих этапов выделяется кислород. (Бактерия Pseudomonas добывает с помощью этого процесса необходимый для дыхания кислород при отсутствии в почве свободного кислорода). Оксид азота (N2
O) и молекулярный азот (N2
) выделяются в атмосферу и, тем самым, исключаются из фондов активного азота. Денитрификация может происходить также чисто химическим путем, без участия микроорганизмов. Например, в кислых почвах происходит реакция:
СО(NH2
)2
+ 2HNO3
® CO2
+ 2N2
+ 3H2
O.
Следует обсудить энергетические взаимоотношения между компонентами круговорота азота, необходимые для функционирования этого круговорота. Ступенчатый процесс разложения белков до нитратов сам служит источником энергии для организмов, осуществляющих это разложение, а обратный процесс требует других источников энергии, таких, как органическое вещество или солнечный свет.
Фиксация азота требует особенно больших затрат энергии, так как много энергии идет на разрыв тройной связи в молекуле азота N2
, чтобы с добавлением водорода из воды превратить ее в две молекулы аммиака (NН3
). Бактерии в клубеньках бобовых расходуют на биофиксацию 1 г азота около 10 г глюкозы (примерно 40 ккал), полученной растением в фотосинтезе, то есть эффективность составляет 10%.
Для фиксации азота необходимы специализированные биохимические механизмы, отсутствующие, по-видимому, у высших растений; лишь прокариоты, безъядерные, самые примитивные организмы, такие как сине-зеленые водоросли, бактерия Azotobacter и др., могут превращать биологически бесполезный газообразный азот в формы, необходимые для построения и поддержания живой протоплазмы. Когда эти микроорганизмы образуют взаимно выгодные ассоциации с высшими растениями, фиксация азота значительно усиливается. Растение предоставляет бактериями подходящее местообитание (т.е. корневые клубеньки, листья), защищает от излишка кислорода, который мешает фиксации, и поставляет им необходимую высококачественную энергию - глюкозу. За это растение получает легкоусваемый фиксированный азот.
Около 80% азота, который, по оценкам, усваивается ежегодно глобальным биотическим сообществом, возвращается в круговорот с суши и из воды, и лишь около 20% необходимого количества - это "новый азот", поступающий из атмосферы с дождем и в результате фиксации. Напротив, из азота, поступившего на поля с удобрениями, очень небольшая часть используется повторно, большая же часть теряется с собираемым урожаем, в результате выщелачивания и денитрификации.
В заключение можно отметить, что благодаря механизмам обратной связи, обеспечивающим саморегуляцию, и хорошей забуференности круговорот азота относительно совершенен, если рассматривать его в масштабе крупных экосистем или всей биосферы. Часть азота из густонаселенных областей суши, пресных вод и мелководных морей уходит в глубоководные океанические отложения и таким образом выключается из круговорота, по крайней мере на время (возможно, на несколько миллионов лет). Эта потеря компенсируется поступлением азота в воздух с вулканическими и промышленными газами. Но в последние годы даже круговорот азота испытывает осложнения, вызываемые антропогенным загрязнением воздуха.
Круговорот серы.
Круговорот серы удачно иллюстрирует связь между воздухом, водой и земной корой, так как сера активно циркулирует в каждом из этих резервуаров и между ними.
Основной доступной формой серы, которая восстанавливается автотрофами и включается в аминокислоты и, соответственно, в белки, является сульфат (SO4
2-
). Первичная продукция обеспечивает включение сульфата в органическое вещество, а экскреция животными служит путем возвращения сульфатов в круговорот. Обмен серы между фондом доступного сульфата и фондом сульфидов железа, находящимися глубоко в почве и в осадках происходит благодаря процессам окисления и восстановления.
Перечислим основные черты биогеохимического круговорота серы:
1. Обширный резервный фонд в почве и отложениях, и меньший - в атмосфере.
2. Центральную роль в быстро обменивающемся фонде играют специализированные микроорганизмы, между которыми существует разделение труда - каждый вид выполняет определенную реакцию окисления или восстановления:
H2
S ® S ® SO4
2-
- бесцветные, зеленые и пурпурные серобактерии;
SO4
2-
® H2
S - (анаэробное восстановление сульфата) - Desulfovibrio;
H2
S ® SO4
2-
- (аэробное окисление сульфида) - тиобациллы;
органическая сера ® SO4
2-
и H2
S - аэробные и анаэробные гетеротрофные микроорганизмы соответственно.
3. Микробная регенерация сульфидов из глубоководных отложений, в результате которой вверх движется газовая фаза (Н2
S).
4. Взаимодействие геохимических и метеорологических процессов (эрозия, осадкообразование, выщелачивание, дождь, абсорбция, десорбция и т.д.) с биологическими процессами продукции и разложения.
5. Взаимодействие воздуха, воды и почвы в регуляции круговорота в глобальном масштабе.
Экосистеме требуется не много серы. Она редко бывает фактором, лимитирующим рост растений и животных. Тем не менее круговорот серы является ключевым в общем процессе продукции и разложения биомассы, ибо он способен регулировать другие круговороты биогенных веществ, в частности фосфора. Когда в осадках образуются сульфиды железа, фосфор из нерастворимой переводится в растворимую форму и становится доступным для организмов. Этот факт несомненно следует учитывать при обсуждении антропогенного воздействия на круговорот серы.
Лекция 12.
Осадочный цикл.
1. Схема осадочного цикла.
2. Круговорот фосфора.
3. Круговорот второстепенных элементов.
4. Круговорот элементов питания.
Схема осадочного цикла.
Большинство элементов и соединений более "привязано" к земле, чем азот, кислород, диоксид углерода и вода; и их круговороты входят в общий осадочный цикл, циркуляция в котором осуществляется путем эрозии, осадкообразования, горообразования и вулканической деятельности, а также биологического переноса. Сообществам биосферы доступны те химические элементы, которые по своей геохимической природе входят обычно в состав пород, обнаруживающихся на поверхности.
Твердые вещества, переносимые по воздуху в виде пыли получили название "осадки"; они могут выпадать на землю с дождями или в виде сухих осадков. Кроме природных веществ (например, образующихся при извержении вулканов, в результате пыльных бурь или лесных пожаров) в состав таких "осадков" входят и вещества, образующиеся в результате деятельности человека. Их сравнительно немного по количеству, но они имеют большое биологическое значение либо из-за своего токсичного, радиоактивного воздействия, либо из-за способности блокировать поступающее солнечное излучение, что может охладить Землю и вызвать климатические изменения, противоположные тем, которые вызываются увеличением содержания парниковых газов.
Прослеживается общая направленность осадочного цикла "вниз". В периоды минимальной геологический активности накопление растворенных или пригодных к использованию минеральных элементов питания происходит на низменностях и в океанах за счет возвышенных районов, хотя скорость этого процесса изменчива. В таких условиях особенно важное значение приобретают местные биологические механизмы возврата, благодаря которым потеря веществ "вниз" не превосходит их поступления из подстилающих пород, иными словами, чем дольше жизненно важные элементы будут оставаться в данной области, вновь и вновь используясь сменяющимися поколениями организмов, тем меньший приток нового материала потребуется извне.
Источники осадочного материала для экосистем верховий невелики. К сожалению, человек часто нарушает этот гомеостаз, обычно не преднамеренно, а просто потому, что не понимает сложности "симбиоза" между живой и неживой материей, для возникновения которого и эволюции, возможно, понадобились тысячелетия. Пример такого "симбиоза" - приспособление речной биоты к передвижению веществ по спирали. Суть этого процесса заключается в том, что водные насекомые, рыбы и другие организмы собирают взвешенные и растворенные вещества, удерживают их, пропускают через пищевую цепь, а более подвижные виды в ходе своего жизненного цикла могут перемещать эти вещества вверх против течения или из реки на водосборный бассейн. Таким образом, действуя совместно, речные животные возвращают в круговорот элементы питания и сокращают их вынос в океан. В высокоширотных районах плотины, препятствующие ходу лососей на нерест, приводят к сокращению численности не только лосося, но и непроходной рыбы, дичи и даже к уменьшению продукции древесины. Удаление из леса больших масс древесины без возврата содержащихся в ней минеральных веществ в почву, что в норме происходит, когда упавшие деревья разлагаются, несомненно, также обедняет нагорья, где запас элементов питания и без того уменьшен веками геологического выщелачивания и эрозии. Нарушение таких биологических механизмов возвращения веществ в круговорот может обеднить целую экосистему на многие годы, так как восстановление обменного фонда минеральных веществ может занять много времени. В таком случае необходимо обдумать какие-то способы возвращения лимитирующих веществ, более эффективные, чем разведение рыбы или лесопосадки.
Внезапно увеличивающийся из-за искусственного усиления эрозии приток элементов питания в низины не обязательно благоприятствует функционированию низинных экосистем, поскольку эти системы могут не успевать ассимилировать элементы питания, проходящие через систему к морю, где, попав в неосвещенные слои воды, они оказываются вне биологического круговорота, по крайней мере на время. С неменьшей вероятностью организмы будут просто задушены потоком ила, грязи и песка или могут быть отравлены токсичными веществами.
Круговорот фосфора.
Фосфор необходим живым организмам в довольно большом количестве, соответствующем примерно 0,1 необходимого количества азота, поскольку он представляет собой один из главных компонентов нуклеиновых кислот, клеточных мембран, систем переноса энергия, костной ткани и дентина. Фосфор имеет важное значение еще и по ряду других причин. Считают, что недостаток фосфора ограничивает продуктивность растений во многих водных местообитаниях и что поступление фосфора в реки и озера со сточными водами и с поверхностным стоком с удобряемых полей стимулирует повышение продуктивности водных местообитаний до нежелательного уровня. Кроме того, изучение круговорота фосфора облегчается тем, что экологи могут без труда измерять его концентрации в экосистеме и следить за его перемещением, используя один из его изотопов в качестве радиоактивной метки.
Круговорот фосфора - пример достаточно простого по структуре осадочного цикла. Фосфор циркулирует, постепенно переходя из органических соединений в фосфаты, которые снова могут использоваться растениями. Растения ассимилируют фосфор в виде фосфат-иона (PO4
3-
) непосредственно из почвы или воды; у животных содержащийся в пище избыточный органический фосфор выводится из организма с мочой в виде фосфатов; некоторые группы бактерий аналогичным образом превращают содержащийся в детрите органический фосфор в фосфат.
Несмотря на относительную простоту круговорота фосфора, на доступность этого элемента для растений оказывают влияние многие факторы среды. При обилии растворенного кислорода фосфор легко образует нерастворимые соединения, которые осаждаются, тем самым изымая фосфор из фонда доступных биогенных элементов. При длительном сохранении таких условий накапливается осадочный фосфор и в конечном счете образуются фосфатные породы, откуда фосфор возвращается в активные фонды экосистемы очень медленно. Кислотность также оказывает влияние на доступность фосфора для растений; фосфаты натрия и кальция относительно плохо растворимы в воде. В щелочной среде фосфат-ионы (PO4
3-
) легко соединяются с натрием или кальцием, образуя нерастворимые соединения. Но в кислой среде фосфат превращается в хорошо растворимую фосфорную кислоту. При промежуточных уровнях кислотности фосфат-ион образует соединения, обладающие промежуточной растворимостью, как это показано на схеме 1:
Схема 1.
| --————————————— Повышение кислотности ——————————————® |
Ионная
форма
|
PO4
3-
®
¯
|
HPO4
2-
®
¯
|
H2
PO4
-
®
¯
|
H3
PO4
|
| Соль |
Na3
PO4
|
CaHPO4
|
NaH2
PO4
|
- |
| Растворимость |
Слабо
растворима
|
Нерастворима |
Растворима |
Очень хорошо растворима |
В лабораторных условиях кислая среда повышает растворимость фосфатов; в условиях экосистемы высокая кислотность снижает доступность фосфора вследствие его реакций с другими минеральными веществами. В кислых средах алюминий, железо и марганец становятся растворимыми и реакционно-способными и образуют химические комплексы, связывающие фосфор и изымающие его таким образом из активного фонда биогенных элементов; именно так происходит извлечение фосфора из почвы в торфяных болотах и в холодных влажных областях, что обуславливает низкую продуктивность этих местообитаний. Фосфор наиболее легкодоступен в узком диапазоне кислотности, в слабокислой среде.
Фосфор поступает в атмосферу в единственной форме - в виде пыли. Поэтому в круговорот фосфора в экосистеме вовлечены только почва и вода. Резервуаром фосфора служит не атмосфера, а горные породы и другие отложения, образовавшиеся в прошлые геологические эпохи. Породы эти постепенно подвергаются эрозии, высвобождая фосфаты в экосистемы, но большое количество фосфатов попадает в море, отлагаясь частично в мелководных осадках, а частично теряясь в глубоководных отложениях. Механизмы возвращения фосфора в круговорот, видимо, не достаточно эффективны и не возмещают потерь. В некоторых районах земного шара сейчас не происходит сколько-нибудь значительного
поднятия отложений, а перенос на сушу рыбы не компенсирует поток фосфора с суши на море. В прошлом морские птицы, по-видимому, играли важную роль в возвращении фосфора в круговорот. Этот перенос фосфора и других веществ из моря на сушу продолжается и сейчас, но, видимо, не столь интенсивно, как в прошлом.
К сожалению, деятельность человека ведет к усиленной потере фосфора, что делает его круговорот менее замкнутым. Хотя человек вылавливает много морской рыбы, полагают, что в год этим способом на сушу возвращается 60000 т элементарного фосфора. Добывается же на удобрения ежегодно 1-2 млн.т. фосфорсодержащих пород; большая часть этого фосфора смывается и выключается из круговорота.
Экологи уделяют большое внимание круговороту фосфора, считая, что его важность сильно возрастет в будущем, так как из всех макроэлементов, т.е. элементов, необходимых для всего живого в больших количествах, фосфор - один из самых редких в смысле его относительного обилия в доступных резервуарах на поверхности Земли.
Круговорот второстепенных элементов.
Второстепенные элементы, подобно жизненно важным, нередко мигрируют между организмами и средой, хотя и не представляют какой-либо известной ценности для организмов. Большинство из них участвуют в общем осадочном цикле, а некоторые поступают в атмосферу. Многие элементы, не относящиеся к биогенным, концентрируются в определенных тканях иногда благодаря химическому сходству с какими-то жизненно важными элементами, хотя такие их концентрации могут оказаться опасными. В настоящее время экологу приходится изучать круговорот целого ряда таких второстепенных элементов (тяжелых металлов, радиоактивных элементов), главным образом потому, что они связаны с деятельностью человека.
Большинство второстепенных элементов в концентрациях, обычных для многих природных экосистем, почти не оказывает влияния на организмы, возможно потому, что организмы к ним адаптировались. Таким образом, миграции этих элементов мало интересовали бы нас, если бы в окружающую среду не слишком часто попадали побочные продукты горнодобывающей промышленности, различных производств химической промышленности и современного сельского хозяйства, продукты, содержащие высокие концентрации тяжелых металлов, ядовитые органические соединения и другие потенциально опасные вещества. Поэтому
в настоящее время важны круговороты почти всех элементов. Даже очень редкий элемент, если он выносится в среду в форме высокотоксичного соединения металла или радиоактивного изотопа, может приобрести важное биологическое значение, так как даже небольшое (с геохимической точки зрения) количество такого вещества способно оказывать выраженный биологический эффект.
Круговорот элементов питания.
Характер круговорота элементов питания в северном умеренном поясе и в тропиках, особенно влажных, различается по ряду важнейших особенностей. В холодных районах большая часть органических веществ и доступных элементов питания все время находится в почве или в отложениях, в тропиках же гораздо больший процент этих веществ содержится в биомассе и циркулирует в пределах органической части экосистемы, чему способствуют различные биологические адаптации, в том числе мутуалистические отношения между микроорганизмами и растениями, удерживающие элементы питания.
Когда вырубают лес в умеренном поясе, в почве остаются элементы питания, сохраняется ее структура, и с площади, освобожденной от леса, в течение многих лет "обычным" способом можно получать урожай, один или несколько раз в год проводя вспашку, высевая однолетние культуры и внося неорганические удобрения. Зимой низкие температуры способствуют удержанию в почве элементов питания и частично уничтожают вредителей и паразитов. Напротив, во влажных тропиках вырубка леса отнимает у земли способность удерживать элементы питания и поддерживать их циркуляцию, так как здесь круглый год сохраняются высокие температуры и повторяются периоды выщелачивающих дождей. Очень часто продуктивность культур быстро падает, и земля забрасывается.
Итак, на севере и круговороты элементов питания, и использование экологических сообществ в большей степени зависят от физических процессов, а на юге - от биологических. Бедная элементами питания тропическая экосистема в естественных условиях способна поддерживать высокую продуктивность благодаря разнообразным механизмам, удерживающим эти элементы и обеспечивающим "прямой круговорот " от растения к растению, в значительной мере минуя почву. Разрушение этих механизмов привозит к снижению продуктивности системы и ее гибели.
Лекция 13.
Пути возвращения веществ в круговорот.
1. Пути возвращения веществ в круговорот.
2. Коэффициент рециркуляции.
Пути возвращения веществ в круговорот.
В связи с вышесказанным, полезно рассмотреть предмет биогеохимии с точки зрения путей возврата веществ в круговорот, поскольку такая рециркуляция воды и элементов питания - жизненно важный процесс в экосистемах, приобретающий большое значение и для человека. Можно выделить пять основных путей возврата в круговорот:
1. Через микробное разложение в детритный комплекс.
2. Через экскременты животных.
3. Прямая передача от растения к растению микроорганизмами-симбионтами.
4. Физическими процессами, в том числе прямого действия солнечной энергии.
5. За счет энергии топлива, например при промышленной фиксации азота.
Согласно классическим представлениям основными агентами регенерации элементов питания считались бактерии и грибы; действительно, в почвах умеренной зоны, где процесс регенерации лучше всего изучен, преобладает путь 1. Там, где мелкие растения, например, трава или фитопланктон, активно выедаются животными, важную роль может играть путь возврата через экскременты животных - путь 2. Непосредственный возврат симбиотическими организмами (путь 3) ) особенно важен в алиготрофных системах с низким содержанием элементов питания и характерен, например для тропиков, где около 40% элементов питания сосредоточено в наземной части экосистемы. Вода возвращается в круговорот в результате прямого воздействия солнечной энергии; и в результате процессов выветривания и эрозии, связанных с потоками воды, вниз по течению элементы осадочных пород выносятся из абиотического резервуара и попадают в биотические циклы (путь 4 ). Человек вмешивается в ход циклов, когда он затрачивает энергию топлива на опреснение морской воды, производство удобрений или на получение из отходов различных металлов (путь 5).
Наконец, следует отметить, что элементы питания могут высвобождаться из остатков растений и
животных и из фекальных комочков даже без участия микроорганизмов, что доказано помещением этих материалов в стерильные условия. Этот способ возврата в круговорот можно назвать автолизом - саморастворением. В водных или влажных местообитаниях, особенно если погибшие растения и животные или неживые частицы малы (т.е. велико отношение их поверхности к объему), еще до начала разложения микроорганизмами может высвободиться от 25 до 75% элементов питания. Автолиз можно считать шестым важным путем возврата, не требующим затрат метаболической энергии.
Возврат в круговорот - это
не "безвозмездная" услуга, на нее почти всегда затачивается энергия. Если источниками энергии для возвращения служат солнечный свет и органическое вещество, то людям не приходится непосредственно затрачивать дорогостоящие виды топлива. Если не нарушать природные механизмы рециркуляции
и не отравлять их, то они в основном реализуют возврат в круговорот воды и элементов
питания. Повторное же использование промышленных материалов - совсем другое дело, оно требует немалых затрат топлива и денежных средств, но когда запасы этих материалов становятся ограниченными, другого выхода нет.
Коэффициент рециркуляции.
Размеры возврата веществ в круговорот в разных экосистемах можно сравнить, рассчитывая коэффициент рециркуляции - соотношение суммарных количеств веществ, циркулирующих между разными блоками экосистемы и общим потоком вещества через всю систему:
 .
.
Здесь CI - коэффициент рециркуляции (возврата); TSTc
- рециркулируемая доля потока вещества через систему и TST - общий поток вещества через систему. Последняя величина определяется как сумма всех поступлений вещества минус изменение его запасов в системе (если это изменение отрицательно), или как суммарный выход вещества плюс изменение его запасов (если оно положительно). Для расчетов потоков и коэффициентов рециркуляция применяют матричный анализ.
Коэффициент рециркуляции
связан с поступлениями каждого элемента извне, его подвижностью и потребностями в нем биоты. Коэффициент возврата бывает низким (0-10%) либо на ранних стадиях развития экосистемы, либо при изобилии ресурсов, либо в случае несущественных
для жизни элементов. СI высок (> 50%) на зрелых стадиях развития экосистемы, при бедности ресурсов и в случае незаменимых для жизни элементов.
Коэффициент рециркуляции не говорит о ее скорости, т.е. скорости, с которой вещества движутся по кругу. Эта скорость значительно выше в тропическом лесу или в теплом океане, чем в тундре или в холодном озере, но удовлетворительные способы ее количественного измерения пока не разработаны.
Повторное использование бумаги.
Бумага - хороший пример рециркуляции в промышленно-городских системах напоминающий рециркуляцию важных элементов в природных системах. Рециркуляция в естественных экосистемах (коэффициент рециркуляции) возрастает при увеличении разнообразия и усложнении биотических компонентов экосистем, или по мере обеднения ресурсов в среде на входе, или по мера накопления в среде на выходе отходов, опасных для жизни в экосистеме, или в результате взаимодействия этих трех процессов вместе.
Пока мы имеем сколько угодно деревьев, бумажных фабрик и неиспользуемой земли для свалки ненужной бумаги, у нас нет стимулов к тому, чтобы затрачивать средства на технику и энергию, необходимые для повторного использования части бумаги, протекающей через город. По мере того, как в пригородах возрастает плотность населения, поднимается стоимость земли, сохранять свалки и места сброса отходов становится все труднее. Среда на входе тоже может оказывать давление, если запасы природной древесины или продукции фабрик неспособны удовлетворить спрос на бумагу. В обоих случаях оказывается "выгодно" подумать о вторичном использовании. Чтобы оно было успешным,
должен существовать рынок сбыта для старых газет и картона, например фабрика по переработке макулатуры. Такая фабрика соответствует механизму экономии энергии путем рециркуляции или диссипативной структуре в природной экосистеме. Другими словами, ко всей системе приходится добавлять новое, эмерджентное свойство, если мы хотим, чтобы экономия, основанная на рециркуляции, была эффективной.
Лекция 14.
Воздействие среды обитания на биоту.
1.Факторы среды.
2.Общие закономерности их действия на живые организмы.
Факторы среды. Общие закономерности их действия на живые организмы.
Среда обитания- этота часть природы, которая окружает живой организм и с которой он непосредственно взаимодействует. Составные части и свойства среды многообразны и изменчивы. Любое живое существо живет в сложном и меняющемся мире, постоянно приспосабливаясь к нему и регулируя свою жизнедеятельность в соответствии с его изменениями. На нашей планете живые организмы освоили четыре основные среды обитания, сильно различающиеся по специфике условий. Водная среда была первой, в которой возникла и распространилась жизнь. В последующем живые организмы овладели наземно-воздушной средой, создали и заселили почву. Четвертой специфической средой жизни стали сами живые организмы, каждый из которых представляет собой целый мир для населяющих его паразитов или симбионтов.
Приспособления организмов к среде носят название адаптаций.Способность к адаптациям - одно из основных свойств жизни вообще, так как обеспечивает самую возможность ее существования, возможность организмов выживать и размножаться. Адаптации проявляются на разных уровнях: от биохимии клеток и поведения отдельных организмов до строения и функционирования сообществ и экологических систем. Адаптации возникают и изменяются в ходе эволюции видов.
Отдельные свойства или элементы среды, воздействующие на организмы, называются экологическими факторами.
Факторы среды многообразны. Они могут быть необходимы или, наоборот, вредны для живых существ, способствовать или препятствовать выживанию и размножению. Экологические факторы имеют разную природу и специфику действия. Экологические факторы делятся на абиотические и биотические, антропогенные.
Абиотические факторы - температура, свет, радиоактивное излучение, давление, влажность воздуха, солевой состав воды, ветер, течения, рельеф местности - это все свойства неживой природы, которые прямо или косвенно влияют на живые организмы.
Биотические факторы - этоформы воздействия живых существ друг на друга. Каждый организм постоянно испытывает на себе прямое или косвенное влияние других существ, вступает в связь с представителями своего вида и других видов, зависит от них и сам оказывает на них воздействие. Окружающий органический мир - составная часть среды каждого живого существа.
Взаимные связи организмов - основа существования биоценозов и популяций; рассмотрение их относится к области синэкологии.
Антропогенные факторы- это формы деятельности человеческого общества, которые приводят к изменению природы как среды обитания других видов или непосредственно сказываются на их жизни. Хотя человек влияет на живую природу через изменение абиотических факторов и биотических связей видов, антропогенную деятельность следует выделять в особую силу, не укладывающуюся в рамки этой классификации. Значение антропогенного влияния на живой мир планеты продолжает стремительно возрастать.
Один и тот же фактор среды имеет различное значение в жизни совместно обитающих организмов разных видов. Например, сильный ветер зимой неблагоприятен для крупных, обитающих открыто животных, но не действует на более мелких, которые укрываются в норах или под снегом. Солевой состав почвы важен для питания растений, но безразличен для большинства наземных животных и т. л.
Некоторые свойства среды остаются относительно постоянными на протяжении длительных периодов времени в эволюции видов. Таковы сила тяготения, солнечная постоянная, солевой состав океана, свойства атмосферы. Большинство экологических факторов - температура, влажность, ветер, осадки, наличие укрытий, пищи, хищники, паразиты, конкуренты и т. д. - очень изменчиво в пространстве и времени. Степень изменчивости каждого из этих факторов зависит от особенностей среды обитания. Например, температура сильно варьирует на поверхности суши, но почти постоянна на дне океана или в глубине пещер. Паразиты млекопитающих живут в условиях избытка пищи, тогда как для свободноживущих хищников ее запасы все время меняются вслед за изменением численности жертв.
Изменения факторов среды во времени могут быть: 1) регулярно-периодическими, меняющими силу воздействия в связи со временем суток или сезоном года или ритмом приливов и отливов в океане; 2) нерегулярными, без четкой периодичности, например изменения погодных условий в разные годы, явления катастрофического характера - бури, ливни, обвалы и т. п.; 3) направленными на протяжении известных, иногда длительных, отрезков времени, например при похолодании или потеплении климата, зарастании водоемов, постоянном выпасе скота на одном и том же участке и т. п.
Экологические факторы среды оказывают на живые организмы различные воздействия, т. е. могут влиять как раздражители, вызывающие приспособительные изменения физиологических и биохимических функций; как ограничители, обусловливающие невозможность существования в данных условиях; как модификаторы, вызывающие анатомические и морфологические изменения организмов; как сигналы, свидетельствующие об изменениях других факторов среды.
Несмотря на большое разнообразие экологических факторов, в характере их воздействия на организмы и в ответных реакциях живых существ можно выявить ряд общих закономерностей.
1.Закон оптимума
. Каждый фактор имеет лишь определенные
пределы положительного влияния на организмы. Результат действия переменного фактора зависит прежде всего от силы его проявления. Как недостаточное, так и избыточное действие фактора отрицательно сказывается на жизнедеятельности особей. Благоприятная сила воздействия называется зоной оптимума экологического фактораили просто оптимумом для организмов данного вида. Чем сильнее отклонения от оптимума, тем больше выражено угнетающее действие данного фактора на организмы (зона пессимума).Максимально и минимально переносимые значения фактора - это критические точки, за пределами которых существование уже невозможно, наступает смерть. Пределы выносливости между критическими точками называют экологической валентностью (диапазоном толерантности)живых существ по отношению к конкретному фактору среды.
Представители разных видов сильно отличаются друг от друга как по положению оптимума, так и по экологической валентности. Так, например, песцы в тундре могут переносить колебания температуры воздуха в диапазоне около 80°С (от +30° до -55°С), тогда как тепловодные рачки Copilia mirabilis выдерживают изменения температуры воды в интервале не более 6°С (от 23° до 29°С). Появление в эволюции узких диапазонов толерантности можно рассматривать как форму специализации, в результате которой большая эффективность достигается в ущерб адаптивности и в сообществе увеличивается разнообразие.
Одна и та же сила проявления фактора может быть оптимальной для одного вида, пессимальной - для другого и выходить за пределы выносливости для третьего.
Широкую экологическую валентность вида по отношению к абиотическим факторам среды обозначают добавлением к названию фактора приставки "эври". Эвритермные виды - выносящие значительные колебания температуры, эврибатные - широкий диапазон давления, эвригалинные - разную степень засоления среды.
Неспособность переносить значительные колебания фактора, или узкая экологическая валентность, характеризуется приставкой "стено" - стенотермные, стенобатные, стеногалинные виды и т. д. В более широком смысле слова виды, для существования которых необходимы строго определенные экологические условия, называют стенобионтными,
а те, которые способны приспосабливаться к разной экологической обстановке - эврибионтными.
2. Неоднозначность действия фактора на разные функции.
Каждый фактор неодинаково влияет на разные функции организма. Оптимум для одних процессов может являться пессимумом для других. Так, температура воздуха от 40° до 45°С у холоднокровных животных сильно увеличивает скорость обменных процессов в организме, но тормозит двигательную активность, и животные впадают в тепловое оцепенение. Для многих рыб температура воды, оптимальная для созревания половых продуктов, неблагоприятна для икрометания, которое происходит в другом температурном интервале.
Жизненный цикл, в котором в определенные периоды организм осуществляет преимущественно те или иные функции (питание, рост, размножение, расселение и т. п.), всегда согласован с сезонными изменениями комплекса факторов среды. Подвижные организмы могут также менять места обитания для успешного осуществления всех своих жизненных функций.
Период размножения является обычно критическим; в этот период многие факторы среды часто становятся лимитирующими. Пределы толерантности для размножающихся особей, семян, яиц, эмбрионов, проростков и личинок обычно уже, чем для неразмножающихся взрослых растений или животных. Так, взрослый кипарис может расти и на сухом нагорье и погруженным в воду, но размножается он только там, где есть влажная, но не заливаемая почва для развития проростков. Многие морские животные могут переносить солоноватую или пресную воду с высоким содержанием хлоридов, поэтому они часто заходят в реки вверх по течению. Но их личинки не могут жить в таких водах, так что вид не может размножаться в реке и не обосновывается здесь постоянно.
3. Изменчивость, вариабельность и разнообразие ответных реакций на действие факторов среды у отдельных особей вида.
Степень выносливости, критические точки, оптимальная и пессимальные зоны отдельных индивидуумов не совпадают. Эта изменчивость определяется как наследственными качествами особей, так и половыми, возрастными и физиологическими различиями. Например, у бабочки мельничной огневки - одного из вредителей муки и зерновых продуктов - критическая минимальная температура для гусениц -7°С, для взрослых форм -22°С, а для яиц -27°С. Мороз в 10°С губит гусениц, но не опасен для имаго и яиц этого вредителя. Следовательно, экологическая валентность вида всегда шире экологической валентности каждой отдельной особи.
4. К каждому из факторов среды виды приспосабливаются относительно независимым путем.
Степень выносливости к какому-нибудь фактору не означает соответствующей экологической валентности вида по отношению к остальным факторам. Например, виды, переносящие широкие изменения температуры, совсем не обязательно должны также быть приспособленными к широким колебаниям влажности или солевого режима. Эвритермные виды могут быть стеногалинными, стенобатными или наоборот. Экологические валентности вида по отношению к разным факторам могут быть очень разнообразными. Это создает чрезвычайное многообразие адаптаций в природе. Набор экологических валентностей по отношению к разным факторам среды составляет экологический спектр вида.
5. Несовпадение экологических спектров отдельных видов.
Каждый вид специфичен по своим экологическим возможностям. Даже у близких по способам адаптации к среде видов существуют различия в отношении к каким-либо отдельным факторам.
6. Взаимодействие факторов.
Оптимальная зона и пределы выносливости организмов по отношению к какому-либо фактору среды могут смещаться в зависимости от того, с какой силой и в каком сочетании действуют одновременно другие факторы. Эта закономерность получила название взаимодействия факторов.Например, жару легче переносить в сухом, а не во влажном воздухе. Угроза замерзания значительно выше при морозе с сильным ветром, чем в безветренную погоду. Таким образом, один и тот же фактор в сочетании с другими оказывает неодинаковое экологическое воздействие. Наоборот, один и тот же экологический результат может быть получен разными путями. Например, увядание растений можно приостановить путем как увеличения количества влаги в почве, так и снижения температуры воздуха, уменьшающего испарение. Создается эффект частичного взаимозамещения факторов.
Вместе с тем взаимная компенсация действия факторов среды имеет определенные пределы, и полностью заменить один из них другим нельзя. Полное отсутствие воды или хотя бы одного из основных элементов минерального питания делает жизнь растения невозможной, несмотря на самые благоприятные сочетания других условий. Крайний дефицит тепла в полярных пустынях нельзя восполнить ни обилием влаги, ни круглосуточной освещенностью.
7. Правило ограничивающих (лимитирующих) факторов.
Факторы среды, наиболее удаляющиеся от оптимума, особенно затрудняют возможность существования вида в данных условиях. Если хотя бы один из экологических факторов приближается или выходит за пределы критических величин, то, несмотря на оптимальное сочетание остальных условий, особям грозит гибель. Такие сильно уклоняющиеся от оптимума факторы приобретают первостепенное значение в жизни вида или отдельных его представителей в каждый конкретный отрезок времени.
Ограничивающие факторы среды определяют географический ареал вида. Природа этих факторов может быть различной. Так, продвижение вида на север может лимитироваться недостатком тепла, в аридные районы - недостатком влаги или слишком высокими температурами. Ограничивающим распространение фактором могут служить и биотические отношения, например занятость территории более сильным конкурентом или недостаток опылителей для растений.
Чтобы определить, сможет ли вид существовать в данном географическом районе, нужно в первую очередь выяснить, не выходят ли какие-либо факторы среды за пределы его экологической валентности, особенно в наиболее уязвимый период развития.
Организмы с широким диапазоном толерантности ко всем факторам обычно наиболее широко распространены.
8. Правило соответствия условий среды генетической предопределенности организма
. Вид организмов может существовать до тех пор и постольку, поскольку окружающая его природная среда соответствует генетическим возможностям приспособления этого вида к ееколебаниям и изменениям. Каждый вид живого возник в определенной среде, в той или иной степени приспособился к ней и дальнейшее его существование возможно лишь в ней или близкой среде. Резкое и быстрое изменение среды жизни может привести к тому, что генетические возможности вида окажутся недостаточными для приспособления к новым условиям.
Лекция 15.
Абиотические факторы среды обитания.
Абиотические факторы, т.е. свойства неживой природы, которые прямо или косвенно влияют на живые организмы, подразделяется на эдафические (почвенные), климатические, топографические и другие физические факторы, в том числе воздействие волн, морских течений и огня.
Климатические факторы внешней среды.
Температура
является наиболее важным климатическим фактором. От нее зависит интенсивность обмена веществ организмов и их географическое распространение.
Любой организм способен жить в пределах определенного диапазона температур. И хотя для разных видов организмов эти интервалы различны (эвритермные и стенотермные организмы), для большинства из них зона оптимальных температур, при которых жизненные функции осуществляются наиболее эффективно, сравнительно невелика. Диапазон температур, в которых может существовать жизнь, составляет примерно 300°С - от -200°С до +100°С. Но большинство видов и большая часть активности приурочены к еще более узкому диапазону температур. Нижний предел выносливости по отношению к температуре лимитируется точкой замерзания внутриклеточной жидкости, при достижении которой клетка обычно физически повреждается и гибнет в результате образования кристаллов льда. Верхний предел накладывает процесс денатурации ферментов. Определенные организмы, особенно в стадии покоя могут существовать, по крайней мере, некоторое время, при очень низких температурах, отдельные виды микроорганизмов, главным образом, бактерии и водоросли способны жить и размножаться при температурах, близких к точке кипения. Верхний предел для бактерий горячих источников составляет 88°С, для сине-зеленых водорослей - 80°С, а для самых устойчивых рыб и насекомых - около 50°С. Между этими крайними точками скорость реакций, контролируемых ферментами, а значит и интенсивность метаболизма удваиваются с повышением температуры на каждые 10°. Поэтому, несмотря на то, что, верхние предельные значения фактора часто оказываются более критическими, чем нижние, многие организмы вблизи верхних пределов диапазона толерантности функционируют более эффективно.
С точки зрения воздействия на живые организмы крайне важна изменчивость температуры. Жизнедеятельность организмов, которые в природе обычно подвергаются воздействию переменных температур, подавляется полностью или частично или замедляется под действием постоянной температуры.
Большинство организмов способно в той или иной степени контролировать температуру своего тела с помощью различных ответных реакций и адаптаций, которые могут смягчать воздействие экстремальных условий и внезапных изменений среды. В водной среде из-за высокой теплоемкости воды не происходит резких изменений температуры, так что в этом отношении условия здесь более стабильны, чем на суше и диапазон толерантности к температуре у водных организмов обычно более узок по сравнению с наземными животными.
Температура, как впрочем и интенсивность света, в большой мере зависит от географической широты, сезона, времени суток и экспозиции склона. Однако встречаются и узколокальные различия в температуре; это в особенности касается микроместообитаний, обладающих собственным микроклиматом. Растительность тоже оказывает некоторое влияние на температуру. Например, иная температура бывает под пологом леса или в меньшей степени внутри отдельных групп растений, а также под листьями отдельного растения.
Таким образом, температура является важным и часто лимитирующим фактором. Температурные ритмы в значительной степени контролируют сезонную и суточную активность растений и животных. Температура часто создает зональность и стратификацию в водных и наземных местообитаниях.
Излучение: свет
представляет собой электромагнитные волны различной длины. Свет необходим для жизни, так как это источник энергии для фотосин
теза, однако есть и другие аспекты его воздействия на живые организмы. Рассматривая эти аспекты, необходимо помнить, что интенсивн
ость света, его качество (длина волны, или цвет) и продолжительн
ость освещения (фотопериод) могут оказывать различное влиян
ие.
Земная атмосфера, включая озоновый слой, селективно, т.е. избирательно по частотным диапазонам, поглощает энергию электромагнитного излучения Солнца и до поверхности Земли доходит в основном излучение с длиной волны от 300 до 10000 нм. Более длинно- и коротковолновое излучение поглощается атмосферой. С увеличением зенитного расстояния Солнца возрастает относительное содержание инфракрасного излучения (от 50 до 72%).
Спектр электромагнитного излучения Солнца весьма широк и его частотные диапазоны различным образом воздействуют на живое вещество. Не известно, имеют ли экологическое значение длинные радиоволны, хотя по мнению некоторых исследователей, эти волны имеют определенное значение для перелетных птиц и других организмов. Роль ультрафиолетового ( с длинами волн менее 390 нм ), видимого ( диапазон волн от 390до 760 нм ) и инфракрасного (с длинами волн больше 760 нм) излучения частично была рассмотрена в разделе “Энергия в экосистемах”.
Интенсивность фотосинтеза несколько варьирует с изменением длины волны света. В наземных экосистемах качественные характеристики солнечного света не настолько изменчивы, чтобы это сильно влияло на интенсивность фотосинтеза, но при прохождении света через воду красная и синяя части спектра отфильтровываются и получающийся зеленоватый свет слабо поглощается хлорофиллом. Однако красные водоросли имеют дополнительные пигменты (фикоэритрины), позволяющие им использовать эту энергию и жить на большей глубине, чем смогли бы зеленые водоросли.
На интен
сивност
ь света влияет угол падения солнечных лучей на земн
ую поверхн
ость; она изменяется в зависимости от широты, сезон
а, времени дня и экспозиции склона. Интенсивность света, падающего на автотрофный ярус, управляет всей экосистемой, влияя на первичную продуктивность. И у наземных, и у водных растений фотосинтез связан с интенсивностью света линейной зависимостью до оптимального уровня светового насыщения, за которым во многих случаях следует снижение интенсивности фотосинтеза при высоких интенсивностях прямого солнечного света. Подавление фотосинтеза объясняется фотоокислением ферментов, что ослабляет синтез, главным образом, синтез белка и увеличивает долю углеводов в продукте; а активное дыхание ведет к расходованию продукта фотосинтеза. У некоторых растений, например, у эвкалипта, фотосинтез не ингибируется прямым солнечным светом. В данном случае происходит компенсация факторов, так как отдельные растения и целые сообщества приспосабливаются к различным интенсивностям света, становясь адаптированными к тени (диатомовые, фитопланктон) или адаптированными к прямому солнечному свету (хлебные злаки).
Продолжительность светового дня (фотопериод), относительно постоянная на экваторе (около 12 ч), в более высоких широтах измен
яется в зависимости от времен
и года. Для растений и животных таких широт характерна ре
акция н
а фотопериод, которая син
хронизирует их активно
сть с времен
ами года. Фотопериод выступает своеобразным “реле времени” или пусковым механизмом, включающим последовательность физиологических процессов, приводящих к росту, цветению многих растений, линьке и накоплению жира, миграции и размножению у птиц и млекопитающих и к наступлению диапаузы у насекомых.
Необходимость света для растений сущест
вен
но влияет на структуру сообществ. Распространение водных растений ограничено поверхностными слоями воды. В наземных экосистемах в процессе конкуренции за свет у растений выработались определенные стратегии, например, быстрый рост в высоту, использование других растений в качестве опоры (у лиан), увеличение поверхности листьев. В лесах это приводит к ярусной структуре сообщества.
Вода
физиологически необходима для любой протоплазмы. С экологической точки зрения она служит лимитирующим фактором как в наземных местообитаниях, так и в водных, где ее количество подвержено сильным колебаниям, или там, где высокая соленость способствует потере воды организмом через осмос. Все живые организмы, в зависимости от потребности их в воде, а следовательно, и по различиям местообитаний, подразделяются на ряд экологических групп: водные или гидрофильные, постоянно живущие в воде; гигрофильные, живущие в очень влажных местообитаниях; мезофильные, отличающиеся умеренной потребностью в воде, и ксерофильные, живущие в сухих местообитаниях. Количество осадков и влажность - основные величины, измеряемые при изучении этого фактора.
Количество осадков
зависит в основном от путей и характера больших перемещений воздушных масс. Например, ветры, дующие с океана, оставляют большую часть влаги на обращенных к океану склонах, в результате чего за горами остается “дождевая тень”, способствующая формированию пустыни. Двигаясь вглубь суши, воздух аккумулирует некоторое количество влаги, и количество осадков опять увеличивается. Пустыни, как правило, расположены за высокими горными хребтами или вдоль тех берегов, где ветры дуют из обширных внутренних сухих районов, а не с океана, например, пустыня Намиб в Юго-Западной Африке. Распределение осадков по временам года - крайне важный лимитирующий фактор для организмов. Условия, создающиеся в результате равномерного распределения осадков, совершенно иные, чем при выпадении осадков в течение одного сезона. В этом случае животным и растениям приходится переносить периоды длительной засухи. Как правило, неравномерное распределение осадков по временам года встречается в тропиках и субтропиках, где нередко хорошо выражены влажный и сухой сезоны. В тропическом поясе сезонный ритм влажности регулирует сезонную активность организмов аналогично сезонному ритму тепла и света в условиях умеренного пояса. В умеренных климатах осадки обычно распределены по сезонам более равномерно (хотя существует много исключений). В таблице 4 приблизительно указаны типы климаксных биотических сообществ, которые можно ожидать при разном годовом количестве осадков, равномерно распределенном по временам года, в умеренных широтах.
Таблица 4
Типы биотических сообществ умеренного пояса в зависимости от годового
количества осадков
| годовое количество осадков (в мм) |
тип сообществ |
| 0 - 250 |
пустыня |
| 250 - 750 |
степь, саванна или редколесье |
| 750 - 1250 |
сухой лес |
| более 1250 |
влажный лес |
В действительности тип биоты определяется не одним количеством осадков, но равновесием между осадками и потенциальной эвапотранспирацией.
Влажность
- параметр, характеризующий содержание водяного пара в воздухе. Абсолютной влажностью называют количество водяного пара в воздухе, выраженное через массу воды на единицу массы воздуха. В связи с зависимостью количества пара, удерживаемого воздухом, от температуры и давления, введено понятие относительной влажности, то есть отношение содержащегося в воздухе пара к насыщающему пару при данных температуре и давлении. Так как в природе существует суточный ритм влажности - повышение ночью и снижение днем, а также ее колебания по вертикали и горизонтали, этот фактор наряду со светом и температурой играет важную роль в регулировании активности организмов. Влажность изменяет эффекты высоты температуры. Например, при условиях влажности близких к критическим, температура оказывает более важное лимитирующее влияние. Аналогично, влажность играет более критическую роль, если температура близка к предельным значениям. Крупные водоемы значительно смягчают климат суши, так как для воды характерна большая скрытая теплота парообразования и таяния. Фактически существуют два основных типа климата: континентальный с крайними значениями температуры и влажности, и морской, которому свойственны менее резкие колебания, что объясняется смягчающим влиянием крупных водоемов.
Доступный запас поверхностной воды зависит от количества осадков в данном районе, но эти величины не всегда совпадают. Так, пользуясь подземными источниками, куда вода поступает из других районов, животные и растения могут получать больше воды, чем ее поступает с осадками. И наоборот, дождевая вода иногда сразу же становится недоступной для организмов.
Газовый состав
атмосферы также является важным климатическим фактором. Примерно 3 -3,5 млрд. лет назад атмосфера содержала азот, аммиак, водород, метан и водяной пар, а свободный кислород в ней отсутствовал. Состав атмосферы в значительной степени определялся вулканическими газами. Из-за отсутствия кислорода не существовало озонового экрана, задерживающего ультрафиолетовое излучение Солнца. С течением времени за счет биотических и абиотических процессов в атмосфере планеты стал накапливаться кислород; началось формирование озонового слоя. Примерно в середине палеозоя потребление кислорода сравнялось с его образованием, в этот период содержание О2
в атмосфере было близко к современному - около 20%. Далее, с середины девона, наблюдаются колебания в содержании кислорода. В конце палеозоя произошло заметное, примерно до 5% современного уровня, снижение содержания кислорода и повышение содержания углекислого газа, сопровождавшееся изменениями климата и, по-видимому, послужившее толчком к обильному “автотрофному” цветению, создавшему запасы ископаемого углеводородного топлива. Затем последовало постепенное возвращение к атмосфере с низким содержанием углекислого газа и высоким - кислорода, после чего отношение О2
/СО2
остается в состоянии так называемого колебательного стационарного равновесия.Химический состав современной атмосферы представлен в таблице 5.
Таблица 5
Концентрация и общее количество газов в атмосфере
| вещество |
концентрация в чистом сухом воздухе на уровне моря
|
общее количество в
атмосфере (109
т)
|
| азот (N2
) |
78,084%а
|
3900000 |
| кислород (O2
) |
20,9476% |
1200000 |
| аргон (Ar) |
0,934% |
67000 |
| водяной пар (Н2
О) |
не учитывается |
14000 |
| диоксид углерода (СО2
) |
346 млн-1
|
2600 |
| неон (Ne) |
18,18 млн-
1
|
65 |
| криптон (Кr) |
1,14 млн-
1
|
17 |
| метан (CH4
) |
2 млн-
1
|
4 |
| гелий (Не) |
5,24 млн-
1
|
4 |
| озон (О3
) летом |
<0,07 млн-
1
|
3 |
| зимой |
<0,02 млн-1
|
| ксенон (Хе) |
0,087 млн-1
|
2 |
| оксид азота (N2
O) |
0,5 млн-1
|
2 |
| оксид углерода (СО) |
следы |
0,6 |
| водород (Н2
) |
0,5 млн-1
|
0,2 |
| аммиак (NН3
) |
следы |
0,02 |
| оксид азота(NO2
) |
<0,02 млн-1
|
0,013 |
| (NO) |
следы |
0,005 |
| диоксид серы (SO2
) |
<1 млн-1
|
0,002 |
| сероводород (H2
S) |
следы
|
0,001 |
| a
По объему. |
Из таблицы следует, что атмосфера состоит в основном из азота, кислорода и относительно меньшего количества аргона и углекислого газа. Все остальные имеющиеся в атмосфере газы содержатся лишь в следовых количествах. Особое значение для биоты имеет относительное содержание кислорода и углекислого газа. Эти биогенные газы оказывают регулирующее действие на процесс фотосинтеза и являются лимитирующими для многих высших растений. У многих растений удается повысить эффективность фотосинтеза, повысив концентрацию углекислого газа; снижение концентрации кислорода также интенсифицирует этот процесс. В опытах на бобовых и многих других растениях экспериментально доказано, что понижение содержания кислорода в воздухе до 5% повышает интенсивность фотосинтеза на 50%. Крайне важную роль играет также азот. Это важнейший биогенный элемент, участвующий в образовании белковых структур организмов.
Большое значение имеют также фи
зические свойства атмосферы: воздух оказывает лишь незначительное
соп
рот
ивление движению и не может служить оп
орой для н
аземн
ых организмов, и это непосредственн
о сказалось на их строении. В то же время некоторые группы животных стали использоват
ь полет как способ п
ередвижения. В атмосфере, так же как в океан
е, постоян
но происходит циркуляц
ия, энергию
для которой поставляет Солнце. Крупномасштабным
результатом циркуляции воздушных масс яв
ляется перераспределен
ие водян
ых паров, так как атмосфера захватывает их в одном месте (где вода испаряется), переносит и отдает в другом месте (где
выпадают осадки), выравнивание температуры , перераспределение других газов, поступающих в атмосферу, в т
ом числе загрязняющих, с последующим вымыванием их из атмосферы с осадками.
Ветер
оказывает лимитирующее воздействие на активность и даже распространение организмов. Ветер способен даже изменять внешний вид растений, особенно в тех местообитаниях, например в альпийских зонах, где лимитирующее воздействие оказывают другие факторы. В открытых горных местообитаниях ветер лимитирует рост растений, приводит к искривлению растений с наветренной стороны. Кроме того, ветер усиливает эвапотранспирацию в условиях низкой влажности.
Большое значение имеют бури, хотя их действие сугубо локально. Ураганы, да и обычные ветры, способны переносить животных и растения на большие расстояния и тем самым изменять состав сообществ.
Барометрическое давление
, по-видимому, не является лимитирующим фактором непосредственного действия, однако оно имеет прямое отношение к погоде и климату, которые оказывают непосредственное лимитирующее воздействие.
Факторы водной среды.
Водные условия создают своеобразную среду обитания организмов, отличающуюся от наземной прежде всего плотностью и вязкостью. Плотность
воды примерно в 800 раз, а вязкость
примерно в 55 раз выше, чем у воздуха. Вместе с плотностью и вязкостью важнейшими физико-химическими свойствами водной среды являются: постоянные перемещения водных масс
в пространстве, способствующие сохранению физических и химических характеристик; температурная стратификация
, то есть изменение температуры по глубине водного объекта; периодические изменения температуры
; прозрачность
воды, определяющая световой режим под ее поверхностью. От прозрачности зависит фотосинтез зеленых и пурпурных водорослей, фитопланктона, высших растений.
Как и в атмосфере важную роль играет газовый состав
водной среды. В водных местообитаниях количество кислорода, углекислого газа и других атмосферных газов, растворенных в воде и потому доступных организмам, сильно варьирует во времени. В водоемах с высоким содержанием органических веществ кислород является лимитирующим фактором первостепенной важности.
Содержание кислорода в поверхностном слое воды зависит, главным образом, от обмена газом между воздухом и водой и варьирует в пределах 6 -12 млн-1
. Растворимость газа уменьшается при повышении температуры и увеличении солености воды. Однако на большой глубине, где отсутствует контакт с атмосферой, этот обмен зависит от расхода кислорода на биологические процессы и пополнения его запасов при смешивании с водой, более богатой кислородом. Как правило, в гидросфере отсутствует равномерное распределение кислорода. Зональность распределения, главным образом, обусловлена ограниченными скоростями смешивания и диффузии кислорода, которые в свою очередь зависят от разности температур, плотности, содержания соли и растворенного кислорода, а также от скорости и направления движения смеси относительно окружающей массы воды. Скорость диффузии растворенного кислорода в массу воды, истощенную по кислороду, является, таким образом, весьма неопределенной переменной величиной, уменьшающейся во времени в зависимости от разницы в содержании кислорода в двух соседних областях, и поэтому является локальной характеристикой.
Всякий раз, когда потребление кислорода превышает восстановление его запасов, возникает более или менее устойчивое состояние истощения, что оказывает заметное влияние на существование живых организмов в смеси воды и отходов, обедненной кислородом. Масштабы этого влияния зависят от относительных скоростей потребления и восстановления кислорода, движения воды и могут быть полностью определены только после нанесенного ущерба.
Когда концентрация растворенного кислорода снижается слишком сильно (менее 2 млн-1
), некоторые виды бактерий начинают получать необходимый им кислород за счет восстановления сульфат-иона. В этом случае водная масса рассматривается как бескислородная система. Для системы становятся характерными совершенно иные химические процессы, чем для обычной воды, содержащей кислород; так, например, доминирующую роль в процессе продукции органического вещества в экосистеме будет играть бактериальный фотосинтез.
Система диоксид углерода - карбонат является одной из наиболее сложных и важных для гидросферы. Она участвует в обмене воздух - поверхностные слои воды, влияет на химию водной системы, биологическую структуру организмов и отложение осадка, содержащего углерод. От этой системы зависит рН среды, что непосредственно влияет на некоторые химические равновесия в данной локальной системе, особенно в отношении ионов, образующих комплексы. Она непосредственно влияет на биологический цикл организмов, в которых углерод используется в процессах развития, гибели и разложения.
В настоящее время принято общее содержание СО2
в гидросфере, равное примерно 4·10-3
моль/л, из них более 80% в форме НСО3
-
. Распределение диоксида углерода неравномерно и частично зависит от биологической активности в данном районе. Подобно кислороду, содержание СО2
в поверхностных слоях воды является функцией его содержания в атмосфере и парциального давления. Однако схемы распределения диоксида углерода и кислорода сильно отличаются друг от друга: наблюдается единая тенденция, к повышению содержания углерода по мере увеличения глубины вследствие оседания продуктов распада погибших организмов из биологически более богатого поверхностного слоя воды.
Усваиваемый углерод является существенной частью питательных веществ и имеет первостепенное значение во всей экологической структуре гидросферы и локализованных районов в особенности. Диоксид углерода играет основную роль в процессе фотосинтеза, и его концентрация, по-видимому, коррелирует с освещенностью или поступлением энергии для таких районов, где реакция фотосинтеза не подавляется другими физико-химическими процессами. Однако высокое содержание СО2
является лимитирующим фактором для животных, так как оно сопровождается низким содержанием кислорода. Например, при слишком высоком содержании свободного углекислого газа в воде погибают многие рыбы.
Кислотность
- концентрация ( активность) ионов водорода - тесно связана с карбонатной системой. Характеризуется величиной рН ( рН = -lg [ H+
] ). Величина рН изменяется в диапазоне 0 £ рН £ 14; при рН = 7 среда нейтральная, при рН<7 - кислая, при рН>7 - щелочная. Если кислотность не приближается к крайним значениям, то сообщества способны компенсировать изменения этого фактора - толерантность сообщества к диапазону рН весьма значительна. Кислотность может служить индикатором скорости общего метаболизма сообщества. В водах с низким рН содержится мало биогенных элементов, поэтому продуктивность здесь крайне мала.
Соленость
- содержание карбонатов, сульфатов, хлоридов и т.д. - является еще одним значимым абиотическим фактором в водных объектах. В пресных водах солей мало, причем около 80% составляют карбонаты. Содержание же минеральных веществ в мировом океане составляет в среднем 35 г
/л
. Организмы открытого океана обычно стеногалинны, тогда как организмы прибрежных солоноватых вод в общем эвригалинны. Концентрация солей в жидкостях тела и тканях большинства морских организмов изотонична концентрации солей в морской воде, так что здесь не возникает проблем с осморегуляцией.
Течение
не только сильно влияет на концентрацию газов и питательных веществ, но и прямо действует как лимитирующий фактор. Многие речные растения и животные морфологически и физиологически особым образом приспособлены к сохранению своего положения в потоке: у них есть вполне определенные пределы толерантности к фактору течения.
Гидростатическое давление
в океане имеет большое значение. С погружением в воду на 10 м давление возрастает на 1 атмосферу. В самой глубокой части океана давление достигает 1000 атмосфер. Многие животные способны переносить резкие колебания давления, особенно, если у них в теле нет свободного воздуха. В противном случае возможно развитие газовой эмболии. Высокие давления, характерные для больших глубин, как правило, угнетают процессы жизнедеятельности.
Эдафические (почвенные) факторы.
Почвой
называют слой вещества, лежащий поверх горных пород земной коры. Русский ученый Докучаев в 1870 году первым рассмотрел почву как динамическую, а не инертную среду. Он доказал, что почва постоянно изменяется и развивается, а в ее активной зоне идут химические, физические и биологические процессы. Почва формируется в результате сложного взаимодействия климата, растений, животных и микроорганизмов. Советский академик почвовед Вильямс дал еще одно определение почвы - это рыхлый поверхностный горизонт суши, способный производить урожай растений. Рост растений зависит от содержания необходимых питательных веществ в почве и от ее структуры.
В состав почвы входят четыре основных структурных компонента: минеральная основа (обычно 50-60% общего состава почвы), органическое вещество (до 10%), воздух (15-25%) и вода (25-30%).
Минеральный скелет почвы
- это неорганический компонент, который образовался из материнской породы в результате ее выветривания.
Свыше 50% минерального состава почвы занимает кремнезем SiO2
, от 1 до 25% приходится на глинозем Al2
О3
, от 1 до 10% - на оксиды железа Fe2
О3
, от 0,1 до 5% - на оксиды магния, калия, фосфора, кальция. Минеральные элементы, образующие вещество почвенного скелета, различны по размерам - от валунов и камней до песчаных крупинок - частиц диаметром 0,02¸2 мм, ила - 0,002¸0,02 мм и мельчайших частиц глины - менее 0,002 мм в диаметре. Их соотношение определяетструктуру почвы. Она имеет большое значение для сельского хозяйства. Глины и суглинки, содержащие примерно равное количество глины и песка, обычно пригодны для роста растений, так как содержат достаточно питательных веществ и способны удерживать влагу. Песчаные почвы быстрее дренируются и теряют питательные вещества из-за выщелачивания, но их выгоднее использовать для получения ранних урожаев, так как их поверхность высыхает весной быстрее, чем у глинистых почв, что приводит к лучшему прогреванию. С увеличением каменистости почвы уменьшается ее способность удерживать воду.
Органическое вещество
почвы
образуется при разложении мертвых организмов, их частей и экскрементов. Не полностью разложившиеся органические остатки называются подстилкой, а конечный продукт разложения - аморфное вещество, в котором уже невозможно распознать первоначальный материал - называется гумусом. В химическом плане это очень сложная смесь изменчивого состава, образованная органическими молекулами различных типов; в основном гумус состоит из фенольных соединений, карбоновых кислот и сложных эфиров жирных кислот. Гумус, подобно глине, находится в коллоидном состоянии; отдельные частицы его прочно прилипают к глине и образуют глино-гумусовый комплекс. Так же как и глина, гумус обладает большой поверхностью частиц и высокой катионообменной способностью. Эта способность особенно важна для почв с низким содержанием глины. Анионы в гумусе - это карбоксильные и фенольные группы. Благодаря своим физическим и химическим свойствам гумус улучшает структуру почвы и ее аэрацию, а также повышает способность удерживать воду и питательные вещества.
Одновременно с процессом гумификации жизненно важные элементы переходят из органических соединений в неорганические, например, азот- в ионы аммония NH4
+
, фосфор - в ортофосфат-ионы H2
PO4
-
, сера - сульфат-ионы SO4
2-
. Этот процесс называется минерализацией.
Почвенный воздух
также как и почвенная вода, находится в порах между частицами почвы. Порозность ( объем пор) возрастает в ряду от глин к суглинкам и пескам. Между почвой и атмосферой происходит свободный газообмен, в результате чего воздух обеих сред имеет сходный состав. Обычно в воздухе почвы из-за дыхания населяющих ее организмов несколько меньше кислорода и больше углекислого газа, чем в атмосферном воздухе. Кислород необходим для корней растений, почвенных животных и организмов-редуцентов, разлагающих органическое вещество на неорганические составляющие. Если идет процесс заболачивания, то почвенный воздух вытесняется водой и условия становятся анаэробными. Почва постепенно становится кислой, так как анаэробные организмы продолжают вырабатывать углекислый газ. Почва, если она небогата основаниями, может стать чрезвычайно кислой, а это наряду с истощением запасов кислорода неблагоприятно воздействует на почвенные микроорганизмы. Длительные анаэробные условия ведут к отмиранию растений.
Почвенные частицы удерживают вокруг себя некоторое количество воды
. Часть ее, называемая гравитационной водой может свободно просачиваться вниз через почву. Это ведет к выщелачиванию, т.е. к вымыванию из почвы различных минеральных веществ, в том числе азота. Гравитационная вода проходит до уровня грунтовых вод, глубина залегания которых колеблется в зависимости от количества выпадающих осадков.
Вода может также удерживаться вокруг отдельных коллоидных частиц в виде тонкой прочной связанной пленки. Эту воду называют гигроскопической. Она адсорбируется за счет водородных связей на поверхностях кварца и глины или на катионах, связанных с глинистыми минералами и гумусом. Эта вода наименее доступна для корней растений, и именно она последней удерживается в очень сухих почвах. Количество гигроскопической воды зависит от содержания в почве коллоидных частиц, поэтому в глинистых почвах ее намного больше - примерно 15% веса почвы, чем в песчанистых - примерно 0,5%. По мере того, как накапливаются слои воды вокруг почвенных частиц, она начинает заполнять сначала узкие поры между этими частицами. а затем распространяется во все более широкие поры. Гигроскопическая вода постепенно переходит в капиллярную, которая удерживается вокруг почвенных частиц силами поверхностного натяжения. Капиллярная вода может подниматься по узким порам и канальцам от уровня грунтовых вод. Растения легко поглощают капиллярную воду, которая играет наибольшую роль в регулярном снабжении их водой. В отличие от гигроскопической эта вода легко испаряется. Тонкоструктурные почвы, например глины, удерживают больше капиллярной воды, чем грубоструктурные, такие, как пески.
Вода необходима всем почвенным организмам. Она поступает в живые клетки путем осмоса. Вода также важна как растворитель для питательных веществ и газов, поглощаемых из водного раствора корнями растений. Она принимает участие в разрушении материнской породы, подстилающей почву, в процессе почвообразовния.
Химические свойства почвы
зависят от содержания минеральных веществ, которые находятся в ней в виде ионов. Некоторые ионы являются для растений ядом, другие - жизненно необходимы. Особое влияние на характеристики почв оказывает концентрация ионов водорода. Флора почв, кислотность которых близка к нейтральному значению ( рН » 7), особенно богата видами. Известковые и засоленные почвы имеют рН=8 ¸9, а торфяные - до 4. На этих почвах развивается специфическая растительность.
В почве обитает множество видов растительных и животныхорганизмов
, влияющих на ее физико-химические характеристики: бактерии, водоросли, грибы или простейшие одноклеточные, черви и членистоногие. Биомасса их в различных почвах равна ( в кг/га): бактерий 1000¸7000, микроскопических грибов - 100¸1000; водорослей 100¸300, членистоногих - 1000; червей 350¸1000.
В почве осуществляются процессы синтеза, биосинтеза, протекают различные химические реакции преобразования веществ, связанные с жизнедеятельностью бактерий. При отсутствии в почве специализированных групп бактерий их роль выполняют почвенные животные, которые переводят крупные растительные остатки в микроскопические частицы и таким образом делают органические вещества доступными для микроорганизмов.
Органические вещества вырабатываются растениями при использовании минеральных солей, солнечной энергии и воды. Таким образом, почва теряет те минеральные вещества, которые растения взяли из нее. В лесах часть питательных веществ вновь возвращается в почву через листопад. Культурные растения за какой-то период времени изымают из почвы значительно больше биогенных веществ, чем возвращают в нее. Обычно потери питательных веществ восполняются внесением минеральных удобрений, которые в основном прямо не могут быть использованы растениями и должны быть трансформированы микроорганизмами в биологически доступную форму. При отсутствии таких микроорганизмов почва теряет плодородие.
Основные биохимические процессы протекают в верхнем слое почвы толщиной до 0,4 метра, так как в нем обитает наибольшее количество микроорганизмов. Одни бактерии участвуют в цикле превращения только одного элемента, другие - в циклах превращения многих элементов. Если бактерии минерализуют органическое вещество - разлагают органическое вещество на неорганические соединения, то простейшие уничтожают избыточное количество бактерий. Дождевые черви, личинки жуков, клещи разрыхляют почву и этим способствуют ее аэрации. Кроме того, они перерабатывают трудно расщепляемые органические вещества.
К абиотическим факторам среды обитания живых организмов относятся также факторы рельефа
(топография)
. Влияние топографии тесно связано с другими абиотическими факторами, так как она может сильно сказываться на местном климате и развитии почвы.
Главным топографическим фактором является высота
. С высотой снижаются средние температуры, увеличивается суточный перепад температур, возрастают количество осадков, скорость ветра и интенсивность радиации, понижаются атмосферное давление и концентрации газов. Все эти факторы влияют на растения и животных. В результате обычным явлением стала вертикальная зональность.
Горные цепи
могут служить климатическими барьерами. Горы служат также барьерами для распространения и миграции организмов и могут играть роль лимитирующего фактора в процессах видообразования.
Еще один топографический фактор - экспозиция склона
. В Северном полушарии склоны, обращенные на юг, получают больше солнечного света, поэтому интенсивность света и температура здесь выше, чем на дне долин и на склонах северной экспозиции. В Южном полушарии имеет место обратная ситуация.
Важным фактором рельефа является также крутизна склона
. Для крутых склонов характерны быстрый дренаж и смывание почв, поэтому здесь почвы маломощные и более сухие, с ксероморфной растительностью. Если уклон превышает 35°, почва и растительность обычно не образуются, а создаются осыпи из рыхлого материала.
Среди абиотических факторов особого внимания заслуживает огонь
или пожар
. В настоящее время экологи пришли к однозначному мнению, что пожар надо рассматривать как один из естественных абиотических факторов наряду с температурой, атмосферными осадками, почвой и т.п.
Пожары как экологический фактор бывают различных типов и оставляют после себя различные последствия. Верховые или дикие пожары, то есть очень интенсивные и не поддающиеся сдерживанию, разрушают всю растительность и всю органику почвы, последствия же низовых пожаров совершенно иные. Верховые пожары оказывают лимитирующее действие на большинство организмов - биотическому сообществу приходится начинать все сначала с того немногого, что осталось, и должно пройти много лет, пока участок снова станет продуктивным. Низовые пожары, напротив, обладают избирательным действием: для одних организмов оно оказываются более лимитирующим, для других - менее лимитирующим фактором и таким образом способствуют развитию организмов с высокой толерантностью к пожарам. Кроме того, небольшие низовые пожары дополняют действие бактерий, разлагая умершие растения и ускоряя превращение минеральных элементов питания в форму, пригодную для использования новыми поколениями растений.
Если низовые пожары случаются регулярно раз в несколько лет, на земле остается мало валежника, это снижает вероятность возгорания крон. В лесах, не горевших более 60 лет, накапливается столько горючей подстилки и отмершей древесины, что при ее воспламенении верховой пожар почти неизбежен.
Растения выработали специальные адаптации к пожару, так же как они сделали по отношению к другим абиотическим факторам. В частности, почки злаков и сосен скрыты от огня в глубине пучков листьев или хвоинок. В периодически выгорающих местообитаниях эти виды растений получают преимущества и огонь способствует их сохранению, избирательно способствуя их процветанию; широколиственные же породы лишены защитных приспособлений от огня, он для них губителен.
Таким образом, пожары поддерживают устойчивость лишь некоторых экосистем. Листопадным и влажным тропическим лесам, равновесие которых складывалось без влияния огня, даже низовой пожар может причинить большой ущерб, разрушив богатый гумусом верхний горизонт почвы, приведя к эрозии и вымыванию из нее биогенных веществ.
Ионизирующее излучение
-излучение с очень высокой энергией - является неотъемлемой характеристикой окружающей среды. Взаимодействуя с веществом, излучение выбивает электроны из атомов и присоединяет их к другим атомам с образованием пар положительных и отрицательных ионов. Ионизация является основной причиной радиационного повреждения цитоплазмы, степень которого пропорциональна числу пар ионов, образовавшихся в поврежденном веществе. Источником естественного, или фонового, излучения служат 1) космические лучи; 2) калий-40 in vivo ( входящий в состав живых тканей); 3) природные радиоактивные изотопы, содержащиеся в горных породах и почве.
Важное экологическое значение имеют a-излучение ( направленный поток ядер атомов гелия 4
2
Не), b- излучение ( быстрые электроны), электромагнитные g- (l = 5·10-11
¸5·10-13 м) и рентгеновское излучение (
l = 5·10-8
¸5·10-12 м). Все перечисленные виды радиации (и корпускулярные, и волновые) в конце концов поглощаются биологическими системами с одинаковыми последствиями: электронные оболочки атомов в клетках деформируются и атомы ионизируются. В итоге биологическое повреждение клеток производят быстродвижущиеся электроны, выбитые из атомов - независимо от типа первоначальной радиации.
Однако по "силе разрушения" в клетке, по плотности выделения энергии на единицу расстояния, пройденного волной или частицей, все перечисленные виды радиации сильно отличаются друг от друга. Так, тяжелые частицы ( a-частицы) создают зону чрезвычайно высокой плотности ионизации, легкие же частицы (электроны или электроны, выбитые рентгеновскими и/или g-лучами) создают зону низкой плотности ионизации, вызывая другие биологические эффекты.
Выделяют два типа биологических повреждений, вызываемых радиацией.
Физический, или "пулеобразный" (I тип). В этом случае выбитые электроны разрушают молекулярные связи непосредственно в структуре, где они были выбиты. Такое прямое воздействие, протекающее очень быстро, служит главной причиной повреждения ДНК в ядрах клеток при облучении, приводя к генетическим мутациям и нарушениям.
Химический, или косвенный (II тип). Здесь ущерб биологической структуре наносят реакционные частицы, которые образовались вдали от этой структуры, но приблизились к ней в результате блужданий. Например, содержащийся в клетке кислород, захватывая выбитые электроны, превращается в ион-радикал О2
-
·. Этот ион-радикал токсичен, так как способен активно окислять фосфолипиды мембран, нарушая их целостность и функционирование.
При первом типе биологического повреждения тяжелая ионизирующая частица (например, a-частица), проходя через ядро клетки, разорвет обе нити ДНК с большей вероятностью, чем легкая частица (b-частица), производящая слабую ионизацию. Однако для второго, косвенного типа воздействия картина представляется обратной. Легкие частицы, создавая при прохождении через клетку низкую локальную концентрацию свободных ион-радикалов или радикалов, более опасны, чем тяжелые частицы. Дело здесь в том, что, чем меньше концентрация радикалов на определенном участке пути, пройденном ионизирующей частицей, тем меньше между радикалами происходит реакций рекомбинации и более длинен путь блужданий самого радикала, а значит, тем вероятнее радикал поразит важную клеточную структуру (например, ДНК или мембрану).
Степень выносливости различных организмов, а также клеток и тканей к действию ионизирующего излучения называют радиочувствительностью. Мерой ее служит значение дозы облучения, вызывающей гибель 50% организмов или клеток, - ЛД50
, которая измеряется в бэрах (бэр). Один бэр - внесистемная единица эквивалентной дозы излучения, равная энергии излучения 100 эрг, поглощенной массой в 1 г с учетом коэффициента качества излучения (1бэр= 0,01 Дж/кг).
Радиочувствительность у различных биологических объектов сильно отличается (табл. 6). Например, доза 200 бэр вызывает гибель зародышей некоторых насекомых на стадии дробления, доза 500 бэр приводит к стерильности некоторых видов насекомых, доза 1000 бэр абсолютно смертельна для млекопитающих. Как показывают данные большей части исследований, наиболее чувствительны к облучению быстро делящиеся клетки.
Таблица 6
Радиочувствительность различных биологических объектов
| Биологический объект |
ЛД50
, бэр
|
| Клетки млекопитающих |
200¸350 |
| Бактерии |
10000¸45000 |
| Дрожжи |
30 000 |
| Инфузории и амебы |
300000¸500000 |
| Взрослые насекомые |
30000¸50000 |
| Мыши и человек |
350¸700 |
| Семена некоторых растений |
100000 |
Воздействие малых доз радиации оценить сложнее, так как они могут вызвать отдаленные генетические и соматические последствия. Например облучение сосны дозой 1 бэр в сутки на протяжении 10 лет вызвало замедление скорости роста, аналогичное однократной дозе 60 бэр. Повышение уровня излучения в среде над фоновым приводит к повышению частоты вредных мутаций.
У высших растений чувствительность к ионизирующему излучению прямо пропорциональна размеру клеточного ядра, а точнее объему хромосом или содержанию ДНК.
У высших животных не обнаружено такой простой зависимости между чувствительностью и строением клеток; для них более важное значение имеет чувствительность отдельных систем органов. Так, млекопитающие очень чувствительны даже к низким дозам радиации вследствие легкой повреждаемости облучением быстро делящейся кроветворной ткани костного мозга. Даже очень низкие уровни хронически действующего ионизирующего излучения могут вызвать в костях и в других чувствительных тканях рост опухолевых клеток, что может проявиться лишь через много лет после облучения.
Для абиотических факторов справедливы все ранее рассмотренные законы воздействия экологических факторов на живые организмы. Знание этих законов позволяет ответить на вопрос: почему в разных регионах планеты сформировались разные экосистемы? Основная причина - своеобразие абиотических условий каждого региона. Популяции концентрируются на определенной территории и не могут быть распространены повсюду с одинаковой плотностью, поскольку имеют ограниченный диапазон толерантности по отношению к факторам окружающей среды и, следовательно, для каждого сочетания абиотических факторов характерны свои виды живых организмов. Множество вариантов сочетаний абиотических факторов и приспособленных к ним видов живых организмов обуславливают разнообразие экосистем на планете.
Лекция 16.
Биотические отношения и роли видов в экосистеме
.
1. Формы биотических отношений.
2. Понятие “экологическая ниша”.
Ареалы распространения и численность организмов каждого вида ограничиваются не только условиями внешней неживой среды, но и их отношениями с организмами других видов. Непосредственное живое окружение организма составляет его биотическую среду
, а факторы этой среды называются биотическими
. Представители каждого вида способны существовать в таком окружении, где связи с другими организмами обеспечивают им нормальные условия жизни.
Выделяют различные формы биотических отношений. Если обозначить положительные результаты отношений для организма знаком “+”, отрицательные результаты знаком “-”, а отсутствие результатов через “0”, то встречающиеся в природе типы взаимоотношений между живыми организмами можно представить в виде таблицы 7.
Таблица 7.
Формы биотических отношений
| Тип взаимодействия |
Результат взаимодействия для вида |
Общий характер взаимодействия |
| А |
В |
| 1. Конкуренция |
- |
- |
Взаимное подавление обоих видов
|
| 2. Хищничество |
+ |
- |
Взаимодействие благоприятно для популяции хищника А и неблагоприятно для популяции жертвы В. Популяция хищника обычно больше, чем популяция добычи |
| 3. Паразитизм |
+ |
- |
Взаимодействие благоприятно для популяции паразита А и неблагоприятно для популяции хозяина В. Популяция паразита обычно меньше, чем популяция хозяина |
| 4. Аменсализм |
- |
0 |
Популяция В подавляет популяцию А, но сама не получает пользы от воздействия и не испытывает отрицательного влияния |
| 5. Комменсализм |
+ |
0 |
Популяция комменсала А получает пользу от объединения, популяции В это объединение безразлично |
| 6. Симбиоз |
+ |
+ |
Взаимодействие, благоприятное для популяций А и В |
| 7. Нейтрализм |
0 |
0 |
Ни одна популяция не влияет на другую |
Эта схематичная классификация дает общее представление о разнообразии биотических отношений. Рассмотрим характерные особенности отношений различных типов.
Конкуренция
является в природе наиболее всеохватывающим типом отношений, при котором две популяции или две особи в борьбе за необходимые для жизни условия воздействуют друг на друга отрицательно.
Конкуренция может быть внутривидовой и межвидовой. Внутривидовая борьба происходит между особями одного и того же вида, межвидовая конкуренция имеет место между особями разных видов. Конкурентное взаимодействие может касаться жизненного пространства, пищи или биогенных элементов, света, места укрытия и многих других жизненно важных факторов. Преимущества в конкурентной борьбе достигаются видами различными способами. При одинаковом доступе к ресурсу общего пользования один вид может иметь преимущество перед другим за счет более интенсивного размножения, потребления большего количества пищи или солнечной энергии, способности лучше защитить себя, адаптироваться к более широкому диапазону температур, освещенности или концентрации определенных вредных веществ.
Межвидовая конкуренция, независимо от того, что лежит в ее основе, может привести либо к установлению равновесия между двумя видами, при котором ни один из видов не будет развиваться столь же успешно, как в отсутствии конкурента, либо к замене популяции одного вида популяцией другого, либо к тому, что один вид вытеснит другой в иное место или же заставит его перейти на использование иной пищи. Установлено, что два одинаковых в экологическом отношении и потребностях вида не могут сосуществовать в одном месте и рано или поздно одинконкурент вытесняет другого. Это так называемый принцип исключения илипринцип Гаузе
.
Популяции некоторых видов живых организмов избегают или снижают конкуренцию переселением в другой регион с приемлемыми для себя условиями, либо переходом на более труднодоступную или трудноусваиваемую пищу, либо сменой времени или места добычи корма, так, например, ястребы питаются днем, совы - ночью; львы охотятся на более крупных животных, а леопарды - на более мелких; для тропических лесов характерна сложившаяся стратификация животных и птиц по ярусам.
Из принципа Гаузе следует, что каждый вид в природе занимает определенное своеобразное место. Оно определяется положением вида в пространстве, выполняемыми им функциями в сообществе и его отношением к абиотическим условиям существования. Это место, занимаемое видом или организмом в экосистеме, называетсяэкологической нишей
. Образно говоря, если местообитание - это как бы адрес организмов данного вида, то экологическая ниша - это профессия, роль организма в месте его обитания.
Вид занимает свою экологическую нишу, чтобы выполнять отвоеванную им у других видов функцию только ему присущим способом, осваивая, таким образом среду обитания и в то же время формируя ее. Природа очень экономна: даже два вида, занимающих одну и ту же экологическую нишу, не могут устойчиво существовать: в конкурентной борьбе один вид вытеснит другой.
Экологическая ниша как функциональное место вида в системе жизни не может долго пустовать - об этом говорит правило обязательного заполнения экологических ниш: пустующая экологическая ниша всегда бывает естественно заполнена. Экологическая ниша, как функциональное место вида в экосистеме позволяет форме, способной выработать новые приспособления, заполнить эту нишу, однако иногда это требует значительного времени. Нередко кажущиеся специалисту пустующие экологические ниши - лишь обман. Поэтому человек должен быть предельно осторожен с выводами о возможности заполнения этих ниш путем акклиматизации. Акклиматизация
- это комплекс мероприятий по вселению вида в новые места обитания, проводимый в целях обогащения естественных или искусственных сообществ полезными для человека организмами. Расцвет акклиматизаторства пришелся на двадцатые - сороковые годы нашего столетия. Однако, по прошествии времени стало очевидно, что либо опыты акклиматизации видов были безуспешны, либо, что хуже, принесли весьма негативные плоды - виды стали вредителями или распространяли опасные заболевания. Иначе и не могло быть: помещенные в чужую экосистему с фактически занятой экологической нишей новые виды вытесняли тех, кто уже выполнял аналогичную работу. Новые виды не соответствовали нуждам экосистемы, иногда не имели врагов и поэтому могли бурно размножаться.
Классическим примером тому является интродукция кроликов в Австралию. В 1859 г, в Австралию из Англии для спортивной охоты завезли кроликов. Природные условия оказались для них благоприятными, а местные хищники - динго - не опасными, так как бегали недостаточно быстро. В результате кролики расплодились настолько, что на обширных территориях уничтожили растительность пастбищ. В некоторых случаях введение в экосистему естественного врага заносного вредителя приносило успех в борьбе с последним, но здесь не все так просто, как кажется на первый взгляд. Завезенный враг совершенно необязательно сосредоточится на истреблении своей привычной добычи. Например, лисы, интродуцированные в Австралию для уничтожения кроликов, нашли в изобилии более легкую добычу - местных сумчатых, не доставляя запланированной жертве особых хлопот
.
Конкурентные отношения отчетливо наблюдаются не только на межвидовом, но и внутривидовом (популяционном) уровне. При росте популяции, когда численность ее особей приближается к насыщению, вступают в действие внутренние физиологические механизмы регуляции: возрастает смертность, снижается плодовитость, возникают стрессовые ситуации, драки и т.п. Изучением этих вопросов занимается популяционная экология.
Конкурентные отношения являются одним из важнейших механизмов формирования видового состава сообществ, пространственного распределения видов популяций и регуляции их численности.
Хищничество и паразитизм
. Поскольку в структуре экосистемы преобладают пищевые взаимодействия, наиболее характерной формой взаимодействия видов в трофических цепях является хищничество, при котором особь одного вида, называемая хищником, питается организмами (или частями организмов) другого вида, называемого жертвой, причем хищник живет отдельно от жертвы. В таких случаях говорят, что два вида вовлечены в отношения "хищник - жертва".
Виды - жертвы выработали целый ряд защитных механизмов. чтобы не стать легкой добычей для хищника: умение быстро бегать или летать, выделение химических веществ с запахом, отпугивающим хищника или даже отравляющих его, обладание толстой кожей или панцирем, защитной окраской или способностью изменять цвет.
У хищников тоже есть несколько способов добычи жертвы. Плотоядные, в отличие от травоядных, обычно вынуждены преследовать и догонять свою жертву. Одни хищники вынуждены быстро бегать, другие достигают своей цели, охотясь стаями, третьи отлавливают преимущественно больных, раненых и неполноценных особей. Еще один путь обеспечения себя животной пищей - это путь, по которому пошел человек - изобретение орудий лова и одомашнивание животных.
Еще один тип взаимодействия видов - паразитизм
. Паразиты питаются за счет другого организма, называемого хозяином, однако в отличие от хищников они живут на хозяине или внутри его организма на протяжении значительной части их жизненного цикла. Паразит использует для своей жизнедеятельности питательные вещества хозяина, тем самым постоянно ослабляя, а нередко убивая его.
Отметим, что когда два вида связаны отношениями паразитизма или хищничества, разрушительный эффект оказывается в общем небольшим в случае, когда популяции развивались вместе в стабильной в течение длительного времени среде. Однако действие окажется совершенно разрушительным, приводящим к полному исчезновению хозяина или жертвы, если два вида стали контактировать недавно или резко изменилась среда обитания. Например, распахивая и возделывая новые территории, а также сознательно или бессознательно перевозя организмы на большие расстояния, человек оказался главной причиной исчезновения многих видов.
От паразитизма отличается аменсализм
, при котором один вид причиняет вред другому, не извлекая при этом для себя никакой пользы. Чаще всего это те случаи, когда причиняемый вред заключается в изменении среды. Так поступает человек, разрушая и загрязняя окружающую среду.
Симбиоз
- это длительное, неразделимое и взаимовыгодное отношение двух или более видов организмов. Например, жвачные животные ( коровы, олени) переваривают клетчатку с помощью бактерий. Стоит только удалить этих симбионтов, и животные погибнут от голода.
Другим вариантом положительных отношений между двумя видами является комменсализм
. Извлекая из хозяина значительную пользу (пища, убежище), виды - комменсалы не приносят ему никакой выгоды или заметного вреда. Например, многочисленные виды насекомых встречаются исключительно в муравейниках, норах грызунов, гнездах птиц, используя их как местообитание с более благоприятным микроклиматом.
Нейтрализм
- это такой тип отношений. при котором ни одна из популяций не оказывает на другую никакого влияния: никак не сказывается на росте его популяций, находящихся в равновесии, и на их плотности. В действительности бывает, однако, довольно трудно при помощи наблюдений и экспериментов в природных условиях убедиться, что два вида абсолютно независимы один от другого.
Обобщая рассмотрение форм биотических отношений, можно сделать следующие выводы:
- отношения между живыми организмами являются одним из основных регуляторов численности и пространственного распределения организмов в природе;
- негативные взаимодействия между организмами проявляются на начальных стадиях развития сообщества или в нарушенных природных условиях; в недавно сформировавшихся или новых ассоциациях вероятность возникновения сильных отрицательных взаимодействий больше, чем в старых ассоциациях;
- в процессе эволюции и развития экосистем обнаруживается тенденция к уменьшению роли отрицательных взаимодействий за счет положительных, повышающих выживание взаимодействующих видов.
Иногда животных, например, многих насекомых, поедающих растения, а также паразитов, хищников рассматривают в качестве естественных врагов тех организмов, за счет которых они существуют. Такой подход в принципе неверен. Паразиты и хищники, зоофаги и фитофаги являются факторами среды по отношению к своим хозяевам, жертвам и т.п. Следовательно, с общеэкологических позиций все онинеобходимы друг другу. В естественных условиях ни один вид не стремится и не может привести к уничтожению другого. Более того, исчезновение какого-либо естественного “врага” из экологической системы может привести к вымиранию того вида, на котором развивается этот “враг”.
Все эти обстоятельства человек должен учитывать при проведении мероприятий по управлению экологическими системами и отдельными популяциями с целью использования их в своих интересах, а также учитывать косвенные последствия, которые могут при этом иметь место.
Лекция 17.
Развитие и эволюция экосистемы.
1. Стратегия развития экосистемы.
2. Концепция климакса.
3.Эволюция экосистемы.
Стратегия развития экосистемы.
Развитие экосистемы,
чаще всего называемое экологическойсукцессией,можно определить по следующим трем параметрам:
1) это упорядоченный процесс развития сообщества, связанный с изменениями во времени видовой структуры и протекающих в сообществе процессов; он определенным образом направлен и, таким образом, предсказуем;
2) сукцессия происходит в результате изменения физической среды под действием сообщества, т.е. сукцессия контролируется сообществом, несмотря на то что физическая среда определяет характер сукцессии, скорость изменения, а часто и устанавливает пределы, до которых может дойти развитие;
3) кульминацией развития является стабилизированная экосистема, в которой на единицу имеющегося потока энергии приходится максимальная биомасса (или высокое содержание информации) и максимальное количество симбиотических связей между организмами.
Последовательность сообществ, сменяющих друг друга в данном районе, называется сукцессией;более или менее переходные сообщества называются по-разному: последовательные стадии, стадии развития,
стадии первых поселенцев,
тогда как терминальная стабилизированная система известна под названием климакса.Замещение видов в сукцессиях вызывается тем, что популяции, стремясь модифицировать окружающую среду, создают условия, благоприятные для других популяций; это продолжается до тех пор, пока не будет достигнуто равновесие между биотическим и абиотическим компонентами.
Cтратегия сукцессии - есть усиление контроля над физической средой (или гомеостаз со средой) в том смысле, что система достигает максимальной защищенности от резких изменений среды.
Изменения, происходящие в основных структурных и функциональных характеристиках развивающейся экосистемы, перечислены в табл.8. Для того чтобы лучше выявить общие тенденции развития, сравниваются ситуации на ранних и поздних стадиях.
Абсолютная величина изменений, скорость изменений и время, необходимое для достижения стационарного состояния, могут варьировать не только в различных климатических и орографических ситуациях, но и для разных признаков экосистемы в одной и той же физической среде. При наличии хороших исходных данных кривые скорости изменений обычно бывают выпуклыми (причем вначале изменения происходят быстрее), но могут быть также двувершинными или циклическими.
Таблица 8.
Модель экологической сукцессии.
Тенденции, которых следуетожидать в развитии экосистем
| Признаки экосистемы |
Развивающиеся стадии |
Зрелые стадии |
1. Отношение P/R (отношение валовой продукции к дыханию)
2. Отношение P/B (отношение валовой продукции к урожаю на корню)
3. Отношение B/E (биомасса, поддержи-ваемая единицей потока энергии)
4. Урожай (чистая продукция сообщества)
5. Пищевые цепи
|
Энергетика сообщества
>1<
Высокое
Низкое
Высокий
Линейные, преимущественно пастбищные
|
~1
Низкое
Высокое
Низкий
Ветвящиеся (пищевые сети), преимущественно детритные
|
6. Общее органическое вещество
7. Неорганические биогенные вещества
8. Видовое разнообразие - компонент многообразия
9. Видовое разнообразие - компонент выравненности
10. Биохимическое разнообразие
11. Ярусность и пространственная гетерогенность (структурное разнообразие)
|
Структура сообщества
Мало
Экстрабиотические
Мало
Мало
Мало
Слабо организованы
|
Много
Интрабиотические
Велико
Велико
Велико
Хорошо организованы
|
12. Специализация по нишам
13. Размеры организма
14. Жизненные циклы
|
Жизненный цикл
Широкая
Небольшие
Короткие и простые
|
Узкая
Крупные
Длинные и сложные
|
15. Круговороты минеральных веществ
16. Скорость обмена биогенных веществ между организмами и средой
17. Роль детрита в регенерации биогенных веществ
|
Круговороты биогенных элементов
Открытые
Высокая
Незначительная
|
Замкнутые
Низкая
Значительная
|
18. Характер роста
19. Продукция
|
Давление отбора
На быстрый рост («г-отбор»)
Количество
|
На регуляцию обратной связью («К-отбор»)
Качество
|
20. Внутренний симбиоз
21. Сохранение биогенных веществ
22. Стабильность (устойчивость к внешним возмущениям)
23. Энтропия
24. Информация
|
Всеобщий гомеостаз
Не развит
С потерями
Низкая
Высокая
Мало
|
Развит
Полное
Высокая
Низкая
Много
|
Биоэнергетика развития экосистемы
Г. Одум и Пинкертон (1955), опираясь на «правило максимума энергии в биологических системах», впервые указали на то, что сукцессия связана с фундаментальным сдвигом потока энергии в сторону увеличения количества энергии, направленного на поддержание системы.
Биоэнергетику экосистемы отражают пункты 1-5 в табл. 8. На ранних стадиях экологической сукцессии уровень первичной продукции, или валовой фотосинтез (P), превосходит уровень дыхания сообщества (R), так что отношение P/Rбольше единицы. Для особого случая при органическом загрязнении типично P/Rменьше единицы. Термин гетеротрофная сукцессиячасто применяют к такой развивающейся экосистеме, где на начальных стадиях R больше, чем P,
напротив, при автотрофной сукцессиина ранних стадиях наблюдается обратное соотношение. Однако, согласно теории, в обоих случаях P/Rприближается к единице по мере развития сукцессии. Иными словами, в зрелых, или «климаксных», экосистемах наблюдается тенденция к равновесию между связанной энергией и энергией, затрачиваемой на поддержание системы (т. е. тотальным дыханием сообщества). Таким образом, коэффициент P/Rможет служить превосходным функциональным показателем относительной зрелости системы.
Пока Pбольше R,
в системе будут накапливаться органическое вещество и биомасса (B) в результате чего отношение P/Bбудет снижаться или соответственно отношения B/P,B/Rили B/E(где E=
P+R)будут увеличиваться. Теоретически, следовательно, урожай на корню, поддерживаемый доступным потоком энергии (E), достигает максимума на зрелых, или климаксных, стадиях. Как следствие этого чистая продукция сообщества, или урожай за годовой цикл, велика на ранних стадиях и мала или равна нулю в зрелой.
Пищевые цепи и пищевые сети
По мере развития экосистемы следует ожидать тонких изменений в структуре пищевых цепей. Относительно простые и линейные связи между организмами, входящими в состав данной пищевой цепи, существуют только на очень ранних стадиях сукцессии. Более того, гетеротрофное использование чистой продукции происходит преимущественно в пастбищных пищевых цепях, т. е. в последовательности растение - травоядное - хищник. На зрелых стадиях, напротив, пищевые цепи превращаются в сложные сети, и при этом большая часть биологического потока энергии следует по детритному пути. При ненарушаемом течении сукцессии имеется достаточно времени для развития все более тесных связей и взаимных адаптаций между растениями и животными, и это ведет к образованию множества механизмов, уменьшающих выедание растений, например к образованию плохо перевариваемых опорных тканей, к развитию механизмов обратной связи между растениями и животными и к увеличению давления хищников на растительноядных животных. Эти и другие механизмы дают биологическому сообществу возможность поддерживать развитую и сложную биологическую структуру, которая смягчает резкие изменения в физической среде.
Разнообразие и сукцессия
Вероятно, наиболее противоречивые из наблюдаемых в сукцессии тенденций касаются разнообразия (табл. 8 (пп. 8-11).
Разнообразие видов, выражаемое отношением числа видов к общей численности или к площади, возрастает на ранних стадиях развития сообщества. О поведении компонента выравненности известно гораздо меньше. Хотя увеличение разнообразия видов, сопровождающееся уменьшением доминирования какого-либо одного вида или небольшой группы видов (т. е. возрастание выравненности или уменьшение избыточности), может восприниматься при сукцессии как в общем вероятное явление, в сообществе происходят другие изменения, которые могут противоречить этой тенденции. В результате увеличения размеров организмов, увеличения продолжительности и сложности жизненных циклов и усиления межвидовой конкуренции, приводящего к конкурентному исключению видов (табл. 8., пп. 12-14), может возникнуть тенденция к уменьшению числа видов, обитающих на данной территории. На стадии «цветения» организмы обычно бывают мелкими и имеют простые жизненные циклы и высокую скорость размножения. Изменения размеров происходят, вероятно, вследствие перемещения биогенных веществ из неорганической в органическую фазу (табл. 8, п. 7) или представляют собой адаптацию к этому переходу. Мелкие размеры дают селективное преимущество в среде, богатой минеральными и органическими компонентами (особенно это относится к автотрофам), так как возрастает отношение поверхности к объему. Однако по мере развития экосистемы неорганические биогенные вещества становятся все более и более связанными в биомассе (т. е. становятся интрабиотическими), так что селективное преимущество переходит к более крупным организмам (либо к более крупным особям данного вида, либо к более крупным видам, либо к тем и другим), обладающим большими возможностями к накоплению и более сложными жизненными циклами. Таким образом, они адаптируются к использованию сезонных или периодических поступлений свободных биогенных веществ и других ресурсов.
Следовательно, возможность дальнейшего возрастания видового разнообразия в сукцессии зависит от того, будет ли увеличение числа потенциальных ниш, происходящее в результате увеличения биомассы, стратификации (табл. 8, п. 9) и других последствий биологической организации, преобладать над противоположными эффектами увеличения размеров и конкуренции.
Большое число видов, выравненность и стратификация - только эти три аспекта разнообразия изменяются в сукцессии. Быть может, еще более важная тенденция - увеличение разнообразия органических веществ, не только тех, которые связаны в биомассе, но и тех, которые выделяются в среду (воздух, почву, воду) как побочные продукты возрастающего метаболизма сообщества. Такое биохимическое разнообразие в пределах отдельных популяций или системы в целом изучено далеко не столь систематически, как видовое разнообразие; здесь трудно делать какие-либо обобщения; тем не менее, вероятно, можно считать, что по мере развития сукцессии увеличивается значение экстраметаболитов как регулятора, стабилизирующего рост экосистемы и ее состав. Такие метаболиты могут в самом деле играть крайне важную роль, предотвращая превышение популяциями равновесной плотности; тем самым они способствуют уменьшению колебаний и установлению стабильности.
Причинно-следственные связи между разнообразием и стабильностью еще неясны, и их необходимо исследовать со многих точек зрения. Если бы можно было показать, что биотическое разнообразие в самом деле повышает физическую стабильность экосистемы или является ее результатом, то это дало бы нам важную руководящую идею для практических мероприятий по охране природы.
Круговорот биогенных элементов
Важной тенденцией в развитии сукцессии является замыкание или «уплотнение» биогеохимических круговоротов основных биогенных элементов, таких, как азот, фосфор и кальций (табл. 8, пп. 15-17). Зрелые системы обладают по сравнению с развивающимися большей способностью захватывать биогенные элементы и сохранять их для круговорота внутри системы; они теряют лишь очень небольшие количества биогенов по сравнению с незрелыми или нарушенными системами.
Давление отбора: количество против качества
Большое количество продукции характеризует молодую экосистему, тогда как высокое качество продукции и регуляция по типу обратной связи - характерные черты зрелой системы (табл. 8, п. 19). Соответственно виды с высокой скоростью размножения и роста имеют больше шансов выжить на ранних стадиях сукцессии. Напротив, при равновесной плотности, характерной для поздних стадий, давление отбора благоприятствует видам с меньшим потенциалом роста, но более высокой способностью к выживанию в условиях конкуренции. Используя обозначения уравнений роста, где r-собственная скорость роста и K-верхняя асимптота, или равновесный размер популяции (гл. Энергия в экосистемах) можно сказать, что «r-отбор» преобладает на ранних этапах развития экосистемы, а «K-отбор» становится преобладающим на поздних стадиях.
Всеобщий гомеостаз
Краткий обзор развития экосистемы выявил сложность взаимодействующих при этом процессов. Хотя можно с полным правом задать вопрос, являются ли все описанные тенденции характерными для всех типов экосистем, вряд ли можно сомневаться в том, что чистый результат деятельности сообщества состоит в усилении симбиоза, сохранении биогенных веществ, повышении стабильности и содержания информации (табл. 8, пп. 20-24). Общая стратегия направлена на достижение такой обширной и разнообразной органической структуры, какая только возможна в границах, установленных доступным притоком энергии и преобладающими физическими условиями существования (почва, вода, климат и т. п.). По мере развития функционального и все более детализированного изучения биотических сообществ сложилось впечатление о важности мутуализма, паразитизма, хищничества, комменсализма и других форм симбиоза. Особенно примечательны в этом отношении тесные связи между неродственными видами, например между кораллами (кишечнополостные) и водорослями или между микоризой и деревьями. По крайней мере во многих случаях биотическая регуляция выедания растительности, плотности популяций и круговорота питательных веществ служит основным механизмом отрицательной обратной связи, который поддерживает стабильность зрелых система предотвращая чрезмерный рост популяций и разрушительные колебания.
Концепция климакса
Терминальным, или стабильным, сообществом развивающейся серии является климаксное сообщество; это самоподдерживающееся сообщество, находящееся в равновесии с физическим местообитанием. В климаксном сообществе в отличие от развивающихся и других нестабильных стадий отсутствует, по-видимому, годовая чистая продукция органического вещества. Иными словами, годовая продукция сообщества и «импорт» уравновешиваются годовым потреблением и «экспортом». Удобно, хотя и несколько произвольно, различать для данной зоны:
1) единственный климатический климакс,
находящийся в равновесии с общими климатическими условиями;
2) различное число эдафических климаксов,изменяющихся в зависимости от локальных условий субстрата.
Первый климакс-это то теоретическое сообщество, к достижению которого направлено все развитие сукцессии в данном районе; оно реализуется там, где физические условия субстрата не столь экстремальны, чтобы изменять воздействие преобладающего климата. В тех местах, где рельеф местности, почва, водоемы, пожары и другие факторы препятствуют развитию климатического климакса, сукцессия заканчивается эдафическим климаксом.
Эти концепции лучше проиллюстрировать на частном примере. В равнинных или слабо холмистых местностях южной части провинции Онтарио, где почва хорошо дренируется, но влажна, терминальная стадия сукцессии представлена кленово-буковым сообществом (доминирующие растения - сахарный клен и бук). Так как этот тип сообщества вновь и вновь можно встретить во всех местах со слабо расчлененным рельефом и умеренным дренажом, считается, что таким должен быть нормальный, немодифицированный климакс в данном районе. Там, где почва более влажная или более сухая, чем в норме (несмотря на воздействие сообществ), мы встречаем несколько отличные типы терминальных сообществ. Еще большие отклонения от климатического климакса наблюдаются на крутых южных склонах, где микроклимат теплее, или в глубоких долинах и на северных склонах, где микроклимат холоднее. Эти климаксы часто похожи на климатические климаксы, находящиеся соответственно южнее или севернее. Таким образом, там, где имеются нормальные климатические и почвенные условия, мы наблюдаем климатический климакс, а при разнообразных локальных комбинациях измененных условий климата и дренажа - различные эдафические климаксы.
Процент площадей, способных поддерживать сообщества климатического климакса, очень различен в разных областях.
Обычно критерием того, является ли данное сообщество климаксным или нет, служит видовой состав. Однако одного этого критерия часто оказывается недостаточно, так как видовой состав может ощутимо изменяться в ответ на сезонные и кратковременные колебания погоды даже в экосистеме, которая в целом остается стабильной. Хорошим показателем может служить отношение P/Rили другие функциональные критерии. Полезны различные статистические соотношения, например отношение пигментов, время оборота (которое больше в климаксе, чем на стадиях развития), коэффициенты вариации, показатели сходства и другие индикаторы стабильности.
Чрезвычайно интересен следующий вопрос: стареет ли зрелая экосистема, подобно тому как стареет организм? Другими словами, развивается ли в экосистеме после длительного периода относительной стабильности, или «возмужалости», вновь несбалансированный метаболизм и становится ли она опять подверженной болезням и другим нарушениям? Некоторые примеры «катастрофических», или «циклических», климаксов подтверждают, вероятно, эту аналогию между развитием особи и сообщества. Примером концепции катастрофического,или циклического, климакса.
могут служить заросли чапарраля в Калифорнии. Здесь результатом биотического развития является кустарниковый климакс, особенно подверженный «катастрофам», в данном случае - естественным пожарам, которые уничтожают не только зрелую растительность, но и продуцируемые ею антибиотики. Затем происходит быстрое развитие травянистой растительности, продолжающееся до тех пор, пока не восстановится доминирующий кустарник. У экосистемы этого типа имеется, следовательно, естественный циклический климакс, контролируемый взаимодействием пожара и антибиотиков.
Другим примером служит еловый лес, в котором при периодических вспышках вредителей погибают крупные зрелые деревья, что способствует энергичному росту молодых деревьев, в значительной части избегающих повреждения. Таким образом ель и вредители образуют естественную самоподдерживающуюся систему.
Применимость теории развития экосистемы к экологии человека
Принципы развития экосистемы имеют важное касательство к взаимоотношениям между человеком и природой, так как стратегия «наибольшей защиты» (т.е. стремление достигнуть максимальной поддержки сложной структуры биомассы), характеризующая экологическое развитие, часто вступает в противоречие со стремлением человека получить «максимум продукции», иными словами получить наивысший возможный урожай.
Осознание экологической основы этого конфликта между человеком и природой -первый шаг в установлении рациональной системы землепользования.
Эволюция экосистемы
Как и при описанном в предыдущих разделах этой лекции развитии в течение коротких промежутков времени, долговременная эволюция экосистемы формируется под влиянием
1) аллогенных (внешних) сил, таких, как геологические и климатические изменения;
2) автогенных (внутренних) процессов, обусловленных активностью живых компонентов экосистемы.
Первые экосистемы, существовавшие 3 млрд. лет назад. были населены мельчайшими анаэробными гетеротрофами, существовавшими за счет органического вещества, синтезируемого в абиотических процессах. Затем последовало возникновение и популяционный взрыв автотрофных водорослей, преобразовавших восстановительную атмосферу в кислородную. На протяжении длительного геологического времени организмы эволюционировали и возникали системы, все возрастающие по сложности и разнообразию, которые 1) могли контролировать атмосферу и 2) содержали в себе все более крупные и высокоорганизованные виды многоклеточных. Считается, что в пределах этого компонента сообщества эволюционные изменения осуществляются главным образом путем естественного отбора, действующего на видовом или более низком уровне.Однако возможно, что естественный отбор на более высоких уровнях также играет важную роль, особенно 1) сопряженная эволюция,т. е. взаимный отбор зависящих друг от друга автотрофов и гетеротрофов, и 2) групповой отбор,
или отбор на уровне сообществ,
который ведет к сохранению признаков, благоприятных для группы в целом, даже если они неблагоприятны для конкретных носителей этих признаков.
Основная литература:
1. Ю.Одум . Экология. М.: Мир, 1986. Т. 1 и 2.
2. Ю.Одум . Основы экологии. М.: Мир, 1975.
Дополнительная рекомендуемая литература
:
1.М.Бигон, Дж. Харпер, К.Р. Таунсенд. Экология. Особи, популяции, сообщества. М.: Мир, 1990. Т. 1 и 2.
2. А.М.Гиляров. Популяционная экология. М.: МГУ, 1990.
3. Д.Ф.Оуэн. Что такое экология? М.: Лесн.пром-сть.1984.
4. Э.Пианка. Эволюционная экология. М.: Мир, 1981.
5. В.Д.Федоров, Т.Г.Гильманов. Экология. М.: МГУ, 1980.
6. Р.Риклефс. Основы общей экологии. М.: Мир, 1979.
7. В.Лархер. Экология растений. М.: Мир, 1978.
8. Э.Макфедьян. Экология животных. М.: Высш. школа, 1963.
9. Э.Макфедьян. Экология животных ( цели и методы). М.: Мир, 1966.
10. Ю.А.Израэль. Экология и контроль состояния природной среды. М.: Гидрометеоиздат, 1984.
11. Н.Н.Моисеев. Экология человечества глазами математика. М.: Молодая гвардия, 1988.
12. Е.И.Захаров, Н.М.Качурин, И.В. Панферова. Основы общей экологии. Тул.политтехн.ин-т. Тула, 1992.
13. Н.М.Чернова, А.М.Былова. Экология. М.: Просвещение, 1988.
14. Р.Дажо. Основы экологии. М.: Мир, 1975.
15. Г.А.Новиков. Основы общей экологии и охраны природы. Л., 1979.
16. В.И.Вернадский. Биосфера. М., 1967.
17. В.И.Вернадский. Живое вещество. М., 1978.
18. Н.Ф.Реймерс. Охраны природы и окружающей человека Среды: Слов.-справ.- М.: Просвещение, 1992.
19. Н.Грин, У.Стаут, Д.Тейлор. Биология. М.: Мир, 1990. Т. 1,2 и 3.
20. Б.Небел. Наука об окружающей среде: Как устроен мир.: В 2-х т. -М.: Мир, 1993.
|

